Wir werden in diesem Kapitel die ersten lebenden Zellen auf der Erde untersuchen. Und wir werden uns fragen: Wie sind sie ins Dasein gekommen? Warum sind sie entstanden? Wie komplex war die erste lebende Zelle auf der Erde? Wie groß war sie? Was hat sie gegessen? Wie hat sie sich vervielfacht? In was für einem Klima hat sie gelebt? Wie viel musste sie über Chemie, Physik, Mathematik und Informationsverarbeitung wissen, um zu leben, um zu überleben und viele zu werden? Beweisen die ältesten Bakterien in den ältesten Ablagerungen der Erde die Hypothese der Evolution, des Neodarwinismus?
Zuerst werden wir uns kurz die Sedimente der Frühen Präkambrium- und Späten Präkambrium-Zeit anschauen. Wir werden uns dann einzellige Organismen anschauen, die an der Grenze des Lebens gedeihen: in heißem und sogar kochendem Wasser. Wir wollen auch versuchen, herauszufinden: Wie werden diese winzigen Geschöpfe viele? Wie benutzen einige von ihnen das Licht der Sonne als Energiequelle? Wie bauen sie ihre Zellwand?. Einige dieser einzelligen Organismen haben sogar einen drehenden Motor. Es ist der kleinste drehende Motor der Welt. Wie macht die Zelle ihn? Und wie funktioniert er?
Seit wann gibt es Leben auf der Erde? Wann ist die erste lebende Zelle auf der Erde entstanden? Wo? Welches sind jetzt die Überreste der ältesten Zellen der Welt? Wie groß und wie komplex war die erste Zelle? Was konnte sie chemisch tun? Was aß sie? Was müsste man alles wissen und können, um die erste Zelle zu erschaffen? Enthält eine lebende Zelle irgendwelche Information, Planung und Design? Wenn ja, warum? Wie viel muss sie über Biochemie und Biophysik wissen, um zu leben? Warum weiß das Bakterium etwas, was ein menschlicher Wissenschaftler zuerst lernen muss?
Beweisen die Sedimente der Frühen Präkambrium-Zeit, dass sich alles Leben auf der Erde von selbst aus der "ersten Zelle" entwickelt hat, dass es von einem gemeinsamen Vorfahren abstammt? Wie sehr unterscheidet sich die heutige Zelle von der ersten Zelle? Hat sich die erste Zelle in höhere Formen des Lebens entwickelt, in Pflanzen, Tiere, Menschenaffen und Menschen? Waren die ersten lebenden Zellen auf der Erde primitiv? Und gibt es jetzt irgendwelche primitiven Zellen? Was haben einige der führenden Wissenschaftler der Welt jetzt darüber herausgefunden?
William Schopf
J. William Schopf arbeitet in der Abteilung Erd- und Weltraum-Wissenschaft, im Institut für Molekulare Biologie, Universität von Kalifornien in Los Angeles, Kalifornien, USA. Er glaubt an die Evolution. Er schreibt in dem Buch Major Events in the History of Life (1992:37) unter "Präkambrische Mikrofossilien":
"Präkambrische Mikrofossilien kommen hauptsächlich in zwei Sorten sedimentärer Ablagerungen vor: In Kieselschiefer (chert). Er besteht aus kleinen Silizium-Körnern (Kieselerde); sie sind dicht ineinander gefügt. Er kommt als das Mineral Quarz (SiO2) vor. Es wurde chemisch abgesetzt und hat mikroskopische Organismen dort versteinert, wo sie gelebt haben (zum Beispiel, in den Schichten der Stromaliten). Und im Schiefer. Dieses Gestein ist entstanden, wo sich Schichten von Ton oder Schlamm abgesetzt haben, zusammen mit Phytoplankton und anderen Resten, am Boden von Seen oder Ozeanbecken. Diese Schichten wurden dann zusammengepresst. Die Mikrofossilien in Kieselschiefer-Ablagerungen sind gewöhnlich verdreht und sonst wie verformt. Doch sie sind meistens nicht zusammengedrückt worden. Sie bestehen aus dreidimensional erhaltenen Zellen mit organischen Wänden. Sie sind völlig eingebettet und durchsetzt vom versteinernden, feinkörnigen Quarz. Die kohlenhaltigen Mikrofossilien in Schiefern haben sich dagegen erhalten, indem sie zusammen gedrückt wurden. Sie wurden zwischen dünnen Schlammschichten (Silt), die sich abgesetzt hatten, zusammengepresst.
Präkambrische Organische Materie
Manchmal haben der Schiefer und der Kieselschiefer die Form der lebenden Zelle nicht bewahrt. Kann man dann trotzdem noch feststellen, ob dort früher einmal einzellige Organismen gelebt haben? Wenn ja, wie?
Prof. J. William Schopf: "Alle lebenden Systeme bestehen aus organischen Verbindungen, aus chemischen Kombinationen der Elemente Kohlenstoff (C), Wasserstoff (H), Sauerstoff (O), Stickstoff (N), und manchmal Schwefel (S) und Phosphor (P). - 'CHONS'. Diese kohlenhaltige organische Materie wird im allgemeinen in altem Gestein, zum Beispiel, in Kohle bewahrt. Das sind die zusammengepressten Reste alter Pflanzenteile. So scheint es vernünftig, anzunehmen, dass die bloße Gegenwart kohlenhaltiger Materie in Präkambrischen Sedimenten eindeutig beweist, dass dort Leben existiert hat. ... Die grundsätzliche Frage, um die es hier geht, lautet: 'Was ist der biologische Ursprung dieser alten kohlenhaltigen Materie? Welche Arten von lebendem System haben sie gemacht?'
"Glücklicherweise stellt die biochemische Organisation lebender Systeme die Mittel bereit, um diese Frage zu beantworten. Organismen sind nicht einfach nur willkürlich angehäufte organische Verbindungen des CHONSP-Typs. Im Gegenteil. Das sind fein abgestimmte organische Systeme, angetrieben von Energie, die aus externen Quellen stammt, (zum Beispiel, Sonnenlicht), Und innen sind sie ein geordnetes, komplexes Netzwerk biochemischer Reaktionen. Diese Reaktionen werden dann durch Proteinverbindungen bewirkt (katalysiert). Das sind Enzyme. Jede biochemische Reaktion wird von einem bestimmten Enzym katalysiert.
"Der erste biochemische Schritt in der anoxischen und oxygenischen Photosynthese, zum Beispiel, ist eine Reaktion, die das Kohlenstoffatom von Kohlendioxyd in eine organische Verbindung einbaut. In photosynthetischen Bakterien und in Cyanobakterien wird diese Reaktion von einem Enzym katalysiert, das als fibulose-Bisphosphat-Carboxylase/Oxygenase bekannt ist oder in Kurzform Rubisco. Wegen der einmaligen Eigenschaften des Rubisco Enzyms bleibt eine isotopische Signatur in seinen Produkten erhalten. Eine Signatur, die man in organischer Materie entziffern kann. Sogar, wenn sie schon 3.500 Milliarden Jahre alt ist". Schopf, J. W. (1992:38, 39).
Wie macht man das?
Prof. J. W. Schopf: "In der Natur gibt es Kohlenstoffatome in drei verschiedenen Formen, oder Isotopen, die sich von einander durch ihre subatomare Struktur unterschieden. Eines dieser Isotope, Kohlenstoff-14 (verkürzt 14C), ist radioaktiv. Weil es instabil ist und im Verlauf der Zeit zerfällt, kann man es nicht in Materialien entdecken, die älter als etwa 50.000 bis 60.000 Jahre sind.
"Die beiden anderen Isotope des Kohlenstoffs, 13C und 12C, sind beide stabil; keines zerfällt im Laufe der Zeit. Deshalb hat es in der ganzen Erdgeschichte zwei Arten von Kohlendioxyd in der Atmosphäre gegeben, 13CO2 und 12CO2. Das Kohlenstoff einbauende Enzym in der Photosynthese, Rubisco, hat die bemerkenswerte Eigenschaft, zwischen diesen beiden Arten von Kohlendioxyd zu unterscheiden. Und es reagiert bevorzugt mit 12CO2. Und deshalb baut es vorwiegend das isotopisch leichtere stabile Kohlenstoff-Isotop, 12C, in die erzeugten organischen Verbindungen ein.
"Deshalb enthalten in den photosynthetischen Bakterien und Cyanobakterien (und auch allen anderen Photoautotrophen), die Produkte der Rubisco-katalysierten Photosynthese etwas mehr 12C, im Vergleich zur Konzentration dieses Isotops im atmosphärischen CO2. So wie Gesundheitsnahrung, 'essen auch die Photoautotrophen leicht' (Sonnenlicht und isotopisch leichten Kohlenstoff)!
"Atmosphärisches Kohlendioxyd spielt auch in anorganischen chemischen Vorgängen eine Rolle: CO2, wenn im Meerwasser gelöst, verwandelt sich zu Bicarbonat (HCO3-). Es kann mit Kalzium reagieren, und bildet dann Kalzium-kohlensaures Salz, CaCO3-), das mineralische Material, aus dem Kalkstein besteht. Als Ergebnis dieser anorganischen Vorgänge, und im Gegensatz zu den 12C-angereicherten Produkten der Photosynthese, enthält der kohlensaure Kohlenstoff in Kalksteinen, im Vergleich zur atmosphärischen CO2 Quelle etwas weniger 12C.
"Der Unterschied in der Menge der Isotope ist zwar gering, die bei diesen verschiedenen Vorgängen entstehen. Man kann sie aber leicht mit einem Massenspektrometer messen. In typischen heutigen Umgebungen wird biologisch erzeugter organischer Kohlenstoff in 12C zu etwa 17 Teilen je Tausend (17 %o), im Verhältnis zum atmosphärischen CO2, angereichert. Der anorganische, kohlensaure Kohlenstoff von Kalkstein in 12C ist um etwa 7%o verarmt. Das ist ein Nettounterschied zwischen den beiden Kohlenstoffarten von 24%o. Die Menge isotopischer Diskriminierung kann variieren und kann von Umweltzuständen abhängen, wie der Menge von CO2 in der Atmosphäre der Erde. Man nimmt an, dass sich seine Konzentration in der Geschichte des Planeten wahrscheinlich ganz beachtlich verändert hat. Trotzdem kann man einen Nettounterschied von etwa 20 %o bis zu mehr als 40 %o zwischen der isotopischen Zusammensetzung biologischen und anorganischen Kohlenstoffes weit in die geologische Vergangenheit zurückverfolgen. Diese isotopische Signatur hat man tatsächlich in Hunderten von Präkambrischen Gesteinsproben entdeckt. Die ältesten sind etwa 3.500 Millionen Jahre alt. Zusammen mit Studien Präkambrischer Stromatoliten und Mikrofossilien liefern diese biogeochemischen Fakten wichtige Beweise für die Existenz und Natur der ersten Lebewesen." Schopf, J. W. (1992:39, 40).
Eine Rekonstruktion der 3.500-Millionen-Jahre alten Umwelt des Apex-Kieselschiefers, ausgestellt im U.S. Naturkundemuseum, Washington, D.C.; K. M. Towe von der Smithonsian Institution hat es entworfen. Aus: J. William Schopf, Cradle of Life (1999:91) Bild 3.11. Vorne im Bild und hinten am Horizont, aktive und ruhende Vulkane. Lava fließt in das flache Meer. Am Ufer wachsen Stromatoliten. Sie sehen aus wie runde Kuchen, die im Wasser sitzen. Einzeller, die ihre Energie vom Sonnenlicht bekommen, haben sie aufgebaut.
Die ältesten bekannten Mikrofossilien
Wo hat man die ältesten Mikrofossilien gefunden? Was zeigen sie uns?
Prof. J. Schopf: "Der beste Beweis, der völlig stichhaltig zu sein scheint - stammt von Mikrofossilien. Man hat besonders schmale, mikroskopische bakterium- oder cyanobacteriumähnliche Fäden gefunden. Man hat sie versteinert in Sedimenten der Swasiland Supergruppe gefunden (in dünnen Schichten des schwarzen Kieselschiefers (chert) der Onderwacht Gruppe), und der Pilbara Supergruppe (in grauem bis schwarzem kohlenhaltigen Kieselschiefer der Warrawoona Gruppe).
"Isotopische Studien zeigen uns: Diese organische Materie in beiden Ablagerungen, die aus kleinen versteinerten Fäden besteht, ist wie Fossilien in einer ähnlichen Art in jüngeren Sedimenten bewahrt worden (in versteinerten Wäldern, zum Beispiel). Sie besteht aus kohlenhaltigem, organischem Material. Einige der Swasiland Fäden scheinen von einer dünnen, hohlen, organischen Hülle umgeben zu sein. Solch eine Hülle kennt man gut in heutigen Bakterien und Cyanobakterien, anders als das, was in Mineralien vorkommt. Diese Zellen sind zu einem Strang zusammengefügt. Sie waren zuerst von einer schleimigen, organischen 'Hülle' umgeben. Und die Pilbara Fäden bestehen eindeutig aus verschiedenen Zellen mit organischen Wänden (Bild 2.5 in seinem Buch), Sie sind in einer Reihe angeordnet, wie die Perlen auf einer Schnur. So etwas gibt es nicht bei Mineralien. Aber die meisten lebenden fadenförmigen Mikroben haben sie. Die Art und Weise, wie diese Zellen angeordnet sind, und dass dort auch Zellen vorkommen, die sich erst zum Teil in einigen der Pilbara-Fäden geteilt haben, zeigt uns: Sie sind durch die gleiche Art Zellteilung entstanden, wie in prokaryotischen Mikroorganismen, die heute leben.
"Man hat sechs verschiedene Arten fossiler Fäden in den Pilbara Kieselschiefern entdeckt, die nach der Größe und der Form ihrer Zellen eingestuft werden, einschließlich ihrer Endzellen, In einigen Fadenarten sind sie abgerundet, oder kegelförmig (Bild ee2.5A). Man hat diese Fäden sorgfältig mit Mikroben verglichen, die heute leben. Sie zeigen uns: Der größte Teil dieser Fossilien ähnelt im Zellaufbau bestimmten Arten lebender Cyanobakterien. (cyano = grünlich-blau).
"Stromatoliten gibt es in den Swasiland und Pilbara Supergruppen. Und die kohlenstoff-isotopische Signatur der organischen Materie ist in diesen beiden Ablagerungen bewahrt worden. Das zeigt auch an, dass es sich um Cyanobakterien handeln könnte. (Aber sie beweisen nicht, dass Cyanobakterien wirklich da waren). Denn die Stromatoliten und die kohlenstoff-isotopischen Werte können auch von photosynthetischen Bakterien stammen. Die Funde zeigen uns, dass Cyanobakterien dort schon vor etwa 3.500 Milliarden Jahren gelebt haben mögen. Und das ist eine faszinierende Möglichkeit: Wenn diese fossilen Mikroben wirklich Cyanobakterien waren, konnten sie bei der Photosynthese Sauerstoff erzeugen. Ihre Gegenwart zeigt an, dass sich Mikroorganismen mit dieser anspruchsvollen biochemischen Fähigkeit schon in dieser frühen Phase der Geschichte des Lebens entwickelt hatten.
"Die Swasiland und Pilbara Fäden sind die ältesten Funde, die wir kennen. Wenn man ihr sehr hohes geologische Alter berücksichtigt, scheinen sie überraschend weit fortgeschritten zu sein. Einige von ihnen kann man nach ihrer Form überhaupt nicht von Mikroben unterscheiden, die heute leben. Und ihre Zellen haben sich anscheinend genauso geteilt wie heute. Sie haben in der gleich Umgebung gelebt. Und sie hatten den gleichen Stoffwechsel, wie die heute lebenden Mikroben, die genauso aussehen wie sie (look-alikes).
"Das flache Wasser des Archaischen Meeres war offensichtlich von einer komplexen, biologisch vielfältigen Gesellschaft von Mikroorganismen bewohnt, die Stromatoliten bauten. Das waren Ökosysteme, die sich selbst erhielten. Dort lebten photoautotrophe Erzeuger und anaerobe, heterotrophe Verbraucher, Mikroben, die die photosynthetisch erzeugte Nahrung wieder verwerteten. Und dort gab es möglicherweise sogar fortgeschrittene, Sauerstoff erzeugende Cyanobakterien." Schopf, J. W. (1992:42-44).
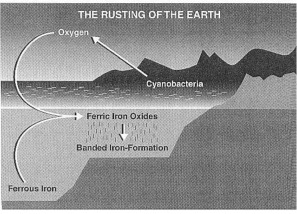
Beweis für gestreifte Eisenformationen
Seit wann gibt es Cyanobakterien auf der Erde, die Sauerstoff erzeugen? Was zeigen uns die gestreiften Eisenformationen des Frühen Präkambriums?
Prof. J. W. Schopf: "Beweise vom sehr veränderten Isua-Gestein zeigen uns, dass Wasser und auch Kohlendioxyd dort schon vor mindestens 3.750 Millionen Jahren vorhanden waren. Weil das Ausgangsmaterial für oxygene Photosynthese schon in dieser entfernten Zeit vorhanden war (und weil Sonnenlicht, das die Photosynthese antreiben muss, schon vorhanden war, als unser Sonnensystem entstand). Deshalb sollten wir uns fragen, ob Sauerstoff erzeugende Cyanobakterien schon in der Isua-Zeit gelebt haben. Das ist gut möglich. Darauf deuten auch die oxydierten Eisenmineralien in der Isua-Formation hin, die dort in den gestreiften Eisenformationen (BIFs) vorkommen (Abb. 2.2).
"Was sind gestreifte Eisenformationen (abgekürzt BIFs)? Und wie passen sie in dieses Bild? ... Kennzeichnend ist: Die Streifen im Gestein bestehen aus nacheinander abgelagerten eisenreichen und eisenarmen Schichten. Die eisenreichen Schichten bestehen aus feinen rostähnlichen Teilchen von Eisenoxiden (besonders aus dem eisenreichen Hematit, Fe2 O3; in einigen Ablagerungen aus dem mineralischen Magnetit, Fe3O4). Die eisenreichen Schichten haben eine auffällig dunkel- bis hellrote Farbe.
"Die Eisenmineralien entstehen, wenn Eisen, durch vulkanische Aktivität entstanden und im Meerwasser gelöst, sich mit molekularem Sauerstoff verbindet. Diese chemische Reaktion ereignet sich gewöhnlich in der oberen Wasserschicht, wo Sauerstoff vorhanden ist. Das Eisenoxyd, das dann entsteht, ist im Meerwasser sehr unlöslich. Deshalb sinkt dann ein feiner Regen winziger rostiger Teilchen auf den Meeresboden." (1992:44).
Woher stammt das Eisen dieser gestreiften Eisenformationen? Und woher kommt der Sauerstoff, der dieses Eisen dann oxydiert?
Prof. J. W. Schopf: "Der größte Teil des atmosphärischen O2 stammt von der Sauerstoff erzeugenden Photosynthese. Dafür gibt es viele Beweise. Bevor die oxygene, cyanobakterielle Photosynthese entstanden ist, kann die Atmosphäre vorher praktisch keinen freien Sauerstoff gehabt haben. Die Umwelt des Planeten muss dann anaerob (ohne freien Sauerstoff) gewesen sein (Abb. 2.2); und der Sauerstoff, den man brauchte, um die BIFs abzusetzen, muss damals sehr knapp gewesen sein.
"Wenn es während der Isua-Zeit viele BIFs gab, und wenn diese Eisenoxid reichen Formationen genauso abgesetzt worden sind, wie später im Präkambrium, dann war das nur möglich, wenn dort schon vor 3.750 Millionen Jahren Sauerstoff erzeugende Photosynthetisierer da waren." (1992:45, 46).
Sauerstoffsenke
Das Cyanobakterium erzeugt freien Sauerstoff. Wenn es dort schon vor etwa 3,5-3,8 Milliarden Jahren gelebt hat, warum gab es dann so wenig freien Sauerstoff in der Atmosphäre des Frühen Präkambriums? Und wie macht das Cyanobakterium diesen freien Sauerstoff?
Prof. J. W. Schopf: "Zusätzlich zu dem Stickstoff, Wasserdampf und Kohlendioxyd, bedeutende Bestandteile der Atmosphäre, geben Vulkane kleinere Mengen anderer Gase ab. Einige von ihnen, wie Wasserstoff, Methan, Kohlenstoffmonoxid und Wasserstoffsulfid, können sich leicht mit molekularem Sauerstoff verbinden. Wasserstoff (H2) verbindet sich, zum Beispiel, mit Sauerstoff. Das heißt, es wird oxydiert, und es entsteht Wasser (H2O); Methan (CH4) und Kohlenstoffmonoxid (CO) wird oxydiert, und erzeugt Kohlenstoffdioxyd (CO2). Und Wasserstoffsulfid (H2S) wird oxydiert, und es entsteht lösliches Sulfat (SO42-). Solche vulkanischen Gase sind deshalb wirksame Sauerstofffresser und dienen als eine der drei Sauerstoffsenken in der frühen Umwelt.
"Sauerstoff, durch Photosynthese erzeugt, wurde auch biologisch verzehrt. Bei der oxygenen Photosynthese wird das Wasser durch die Energie des Lichts (H2O) in Wasserstoff und Sauerstoff gespalten. Der Wasserstoff des Wassers verbindet sich mit Kohlendioxyd, um organische Materie zu erzeugen (gewöhnlich in Kurzschrift geschrieben als Zucker, 'CH2O'). Und der Sauerstoff von diesem Wasser entweicht dann in die Umgebung, als unbenutztes Nebenprodukt der Photosynthese. Die netto chemische Reaktion der cyanobakteriellen Photosynthese ist daher
(Licht-Energie) + Wasser-Kohlendioxyd > organische Materie +Sauerstoff.
Aerobe Atmung
Was passiert während der aeroben Atmung, wenn die Organismen atmen?
Prof. J. W. Schopf: "Dies ist das genaue Gegenteil der biochemischen Reaktion, die während der aeroben Atmung vor sich geht. Alle aeroben Organismen, fakultative und obligate aerobe, und auch die Pflanzen, Cyanobakterien und aerobe Bakterien 'atmen'. Wenn die Organismen 'atmen', oxydieren sie organische Materie, um Energie freizusetzen, indem sie folgende netto chemische Reaktion ausführen:
Organische Materie + Sauerstoff > Wasser + Kohlendioxyd + (zellulare Energie).
So haben die Mikroorganismen, die aerob atmen können, eine zweite Sauerstoffsenke geschaffen. Die frühesten Mikroben waren zweifellos fakultativ und atmeten aerob, wenn Sauerstoff da war. Aber sie schalteten auf anaerobe Gärung um, wenn dort zu wenig Sauerstoff vorhanden war (zum Beispiel, wenn Vulkane aktiv waren). Die vulkanischen Gase haben dann den Sauerstoff verzehrt. Fakultative Aeroben, wirksame Sauerstofffresser, saugten dann den photosynthetisch erzeugten Sauerstoff auf, bevor er sich in der Atmosphäre anreichern konnte." (1992:47, 48).
Gelöstes Eisen
Wie verschluckt das gelöste Eisen die freien Sauerstoffmoleküle?
Prof. J. W. Schopf: "Die dritte Sauerstoffsenke kam vom gelösten Eisen, das sich überall in den Meeresbecken der Welt in der frühen Zeit der Erde angesammelt hatte. Wenn das Eisen in Gebiete gespült wurde, wo photosynthetisch erzeugter Sauerstoff vorhanden war, dann verband sich der Sauerstoff mit dem Eisen. So entstanden unlösliche Eisenoxidmineralien. Die mineralischen Teilchen sanken als feiner, rostiger Regen auf den Meeresboden; und die BIFs wurden abgesetzt. Dabei wurde der photosynthetisch erzeugte Sauerstoff aus den Meeren entfernt und wurde für immer als Rost begraben." (1992:48).
Der Ozean begann zu rosten, als Eisen aus der Tiefe des Meeres oxidierte (verrostete) und die gestreiften (= geschichteten) Eisenformationen bildete. Aus J. W. Schopf, Cradle of Life (1999:172) Bild 6.2. Die Bakterie gewinnt ihre Energie durch das Sonnenlicht, wie auch heute noch. Sie erzeugt dabei freien Sauerstoff, indem sie das Wassermolekül aufspaltet. Dieser freie Sauerstoff verbindet sich dann mit dem noch nicht oxidierten Eisen, das im Wasser gelöst ist. Es wird dann zu Ferri-Eisen (Eisen III). Das heißt, mit Sauerstoff verbundenes Eisen, zu Rost. Dieses oxidierte Eisen (dieser Rost) sinkt dann auf den Meeresboden und bildet die gestreifte Eisenformation (BIF).
Prokaryoten: entwickelt?
Hat sich die prokaryotische Zelle in den fast zwei Milliarden Jahren der Proterozoischen Zeit entwickelt? Mit anderen Worten: Stammen die heutigen Bakterien von einer einfachen, primitiven Urzelle ab? Und haben sich aus dieser ersten Zelle des Frühen Präkambriums dann Pflanzen, Tiere und Menschen entwickelt?
Prof. J. W. Schopf: "Die Antwort lautet: Darüber kann man viel sagen: jetzt weiß man schon recht viel darüber. Überraschenderweise, ja sehr seltsam ist dies: Besonders die Cyanobakterien, und vielleicht auch alle Prokaryoten, scheinen sich von der frühen Proterozoischen Zeit an bis jetzt, kaum entwickelt zu haben! Das ist wirklich seltsam. ... Von ihrer Form her kann man die heute lebenden und die Proterozoischen Cyanobakterien praktisch nicht voneinander unterscheiden. Die Abbildung 2.8 (in seinem Buch) zeigt uns nur vier von einer großen Anzahl von Beispielen. Sie zeigen uns, wie genau sich die heutigen und fossilen Einzeller ähneln.
"Vergleichen Sie, zum Beispiel, das lebende Cyanobakterium Lynbya, in Abb. 2.8A, mit dem fossilen Paleolyngbya, in Abb. 2.8B: Beide sind etwa gleich groß; beide setzen sich aus einzelnen Reihen scheibenförmiger Zellen zusammen; beide haben an den Enden abgerundete Zellen; beide stecken in röhrenförmigen, organischen Scheiden. Die fossilen Fäden (Abb. 2.8B) sind fast eine Milliarde Jahre alt. Aber wenn sie heute leben würden, würde man sie bestimmt in die gleiche Gattung stellen, und vielleicht sogar in die gleiche Spezies, wie die modernen Fäden.
"Oder vergleichen wir das lebende schraubenförmige Cyanobakterium Spirulina (Abb. 2.8C) mit seinem 850 Millionen Jahre alten fossilen Doppelgänger (Abb. 2.8D); oder die in einer Scheide steckende, vierzellige Kolonie von modernen Gloeocapsa, die in Abb. 2.8E gezeigt wird, mit der besonders ähnlichen 1.550 Millionen Jahre alten Gloeocapsa-ähnlichen Kolonie, die in Abb. 2.8F gezeigt wird; oder vergleichen wir die vielzellige Kolonie lebender Entophysalis (Abb. 2.8G) mit ihrem fossilen Gegenstück, dem 2.150 Millionen Jahre alten kolonialen Eoentophysalis (Abb. 2.8H).
"Es gibt noch weitere Beweise. Die Spanne des Durchmessers und das Muster der Größenverteilung der röhrenförmigen Scheiden lebender Cyanobakterien und von vergleichbaren fossilen prokaryotischen Fäden ist praktisch gleich (Abb. 2.9). Außerdem gibt es keinen erkennbaren Trend, kein offensichtliches Muster evolutionärer Änderung, in der Größe, weder bei den fossilen röhrenförmiger Scheiden, noch bei den fossilen zellularen Fäden während der ganzen Proterozoischen Zeit, eine Zeit, die etwa vier Male länger ist, als das ganze Phanerozoikum!" (1992:50, 52).
Was schließen Sie daraus?
Prof. J. W. Schopf: "Der Schluss scheint unausweichlich zu sein. Ganz gleich, ob die Organismen faserig sind oder kugelförmig, ob sie gerade oder schraubenförmig sind, ob sie einzeln vorkommen oder in vielzelligen Kolonien, ganz gleich, wie groß die Zelle sein mag, oder wie die Zelle geformt ist, oder wie ihre äußere Scheide beschaffen ist, die Form der Cyanobakterien hat sich nur wenig, wenn überhaupt, seit dem Anfang der Proterozoischen Zeit verändert."
"Die Größe, Form, und zellulare Organisation der Cyanobakterien ist in mehr als einer Milliarde, und in einigen Fällen, in mehr als zwei Milliarden Jahren unverändert geblieben. ... Außer in der Form, gibt es auch noch viele andere offensichtliche Ähnlichkeiten zwischen lebenden und Proterozoischen Cyanobakterien (zum Beispiel, die Umwelt, in der sie vorkommen, ihre Fähigkeit, Stromatoliten zu bilden; ihre Konzentration auf der Wuchsoberfläche in mikrobiellen Gesellschaften, und die besonderen Arten von Cyanobakterien, den Familien und den Gattungen, die in diesen Gesellschaften vorkommen). Wenn man alles berücksichtigt, scheint es recht wahrscheinlich, dass sich die grundlegende Natur dieser Prokaryoten wenig, wenn überhaupt, seit dem Anfang der proterozoischen Zeit entwickelt hat." (1992:53).
Was schließen Sie jetzt aus Ihren Ergebnissen? Seit wann gibt es Leben auf der Erde?
J. W. Schopf und C. Klein: "Die Zellgröße der meisten fossiler Trichome scheint mehr darauf hinzudeuten, dass es sich hier um cyanobakterielle, und nicht um bakterielle Zellen handelt. ... Wenn man sie mit den heutigen Mikroorganismen vergleicht, scheint es sich bei den meisten faserigen Arten, die man jetzt im Warrawoona Kieselschiefer gefunden hat, um Cyanaobakterien zu handeln, statt um Bakterien. Einige der heutigen Cyanobakterien können zwar auf anoxygene Photosynthese umschalten. Doch die Fähigkeit, Sauerstoff erzeugende Photoautotrophie auszuführen, ist eine universale Eigenschaft dieser Gruppe. Aufgrund dieser Beobachtungen scheint es vernünftig zu sein, wenn man daraus schließt, dass Cyanobakterien, und damit Sauerstoff erzeugende Photosynthetisierer, wahrscheinlich im Lebensraum von Warrawoona vorhanden waren. Und dass sie diese recht fortschrittliche physiologische Evolution schon vor mindestens 3.500 Millionen Jahren erreicht haben.
"Es ist offensichtlich, dass es schon vor ~ 3.500 Millionen Jahren mikrobielle Populationen gegeben hat, in verschiedenen Formen, und möglicherweise physiologisch fortgeschritten. Diese Archaischen Mikrofossilien, sowohl faserig, als auch kolonial, sind in morphologischem Detail den heutigen Prokaryoten besonders ähnlich. Diese Ähnlichkeit findet man sogar in den Mechanismen der Zellteilung." Schopf und Klein (1992:38, 39).
"Die bekannten stromatolitischen, mikrobiellen und kohlenstoff-isotopischen fossilen Funde scheinen eindeutig zu beweisen, dass dort schon vor 3,5 bis 3,4 Milliarden Jahren photoautotrophe Prokaryoten (nämlich, photosynthetische Bakterien und, möglicherweise, O2 erzeugende Cyanobakterien) gelebt haben. ... Anaerobe Bakterien hat es schon vor ~3,5 Milliarden Jahren gegeben. Der reduzierte Kohlenstoff in der sehr veränderten ~3,8 Milliarden Jahre alten Isua Supracrustal Group in Südwest Grönland (Schidlowski et al. 1983) - wenn nicht ein Produkt lebloser Prozesse -, scheint anzuzeigen, dass solche Mikroben noch früher in geologischer Zeit gelebt haben."
"Archaische Stromatoliten kennt man aus den frühesten Sedimenten der ~3,5 Milliarden Jahre alten Warrawoona Gruppe von West Australien. Das deutet stark darauf hin, dass dort photosynthetische Bakterien gelebt haben (und deshalb auch O2 erzeugende Cyanobakterien). Wenigstens die schmaleren faserigen Mikrofossilien in einigen Frühen Archaischen stromatolit-ähnlichen dünnen Schichten (z.B., diejenigen in Kieselschiefer-Sedimenten der ~3,4 Milliarden Jahren alten Ondervacht Gruppe von Südafrika) ähneln morphologisch den heutigen photosynthetischen Bakterien (Chloroflexus). Und kohlenstoff-isotopische Verhältnisse im Kerogen in den Sedimenten der Warrawoona und Onverwacht Gruppe stimmen mit der photoautotrophen Kohlenstoff-Fixierung überein (z.B., bei photosynthetischen Bakterien und/oder Cyanobakterien).
"Der Schluss, den wir daraus ziehen müssen, scheint klar zu sein: Anoxygene photoautotrophe Bakterien sind schon vor mindestens ~3,5 Milliarden Jahren entstanden. Das graphitische kohlenhaltige Material in den ~3,8 Milliarden Jahre alten Sedimenten der Isua Supracrustal Group scheint anzuzeigen, dass solche Organismen noch früher da waren. ... Die altumweltlichen, stromatolitischen, mikrofossilen, und kohlenstoff-isotopischen Fakten, die wir jetzt aus der Zeit vor 3,5 bis 3,3 Milliarden Jahren kennen (sowie die gestreiften Eisenformationen und der reduzierte Kohlenstoff in diesen Sedimenten und in Sedimenten, die bis zu ~3,8 Milliarden Jahre alt sind), beweisen alle, dass im Frühen Archaikum Sauerstoff erzeugende Cyanobakterien gelebt haben. ... Cyanobakterien waren schon im Späten Archaikum vorhanden, und es kann gut sein, dass sie dort schon im Frühen Archaikum gelebt haben." Schopf und Klein (1992:587-590).
![]()
Domförmige und säulenförmige Stromatoliten wachsen heute an der Shark Bay, Westaustralien (A). Sie sehen fast genauso aus wie die fossilen Stromatoliten, die Milliarden von Jahren alt sind (B), aus der 2.300-Millionen-Jahre alten Transvaal Dolomite Formation der Kap Provinz, Südafrika. Der geologische Hammer in (A) rechts in der Mitte, zeigt den Maßstab der beiden Fotos. Aus: J. W. Schopf, Cradle of Life (1999) Bild 7.7. Das zeigt uns: Die Cyanobakterien und ihre Gefährten, welche diese Stromatoliten Schicht um Schicht, in den letzten 2.300 Millionen Jahren aufgebaut haben, haben sich überhaupt nicht verändert. Sie wissen nichts von Evolution.
Auch biochemisch ähnlich?
Viele der beinahe 300 prokaryotischen Ordnung proterozoischer mikrobieller Arten gleichen in ihrer Form bestimmten, modernen Mikroorganismen. Viele der fossilen cyanobakteriellen Arten, die man jetzt kennt, scheinen während eines großen Teils der Proterozoischen Zeit gelebt zu haben (Schopf und Klein 1992:596). Bedeutet dies dann, dass sie sich auch biochemisch ähneln?
Schopf und Klein: "Über die Genetik kann man nur wenig sagen. Doch man hat festgestellt: (i) Die modernen und fossilen Arten ähneln einander so sehr, dass sie auch fundamentale Ähnlichkeiten in den genetischen und Entwicklungs-Kontrollen widerspiegeln, die ihre Formen beherrschen; (ii) Direkte fossile Beweise scheinen zu zeigen, dass die modernen Mechanismen der prokaryotischen (und der cyanobakteriellen) Zellteilung schon vor 3,4 Milliarden Jahren vorhanden waren; und (iii) praktisch der ganze Bereich morphologischer Produkte solcher Zellteilung, die man bei den heutigen Cyanobakterien findet, hatten auch schon die Proterozoischen Arten. Die Form der mittleren (z.B., scheibenförmigen, quadratischen, fassförmigen, länglichen) oder die Zellen am Ende (kugelförmig, kegelförmig, gerundet, mit stumpfem Ende) von cyanobakteriellen Trichomen, von den Zellformen in nicht-faserigen Kolonien (kugelförmig oder elipsenförmig), oder von der Organisation der Zellen, die cyanobakterielle koloniale Haufen bilden (z.B., die nicht in einer Reihe angeordnet sind, in nicht-verzweigten Trichomen vorkommend; unregelmäßige Kolonien; und in Reihen und als Säulen, X-förmig, würfelförmig, rosettenförmig, und ähnlichen sehr geordneten Kolonien).
"Die fossilen Funde beweisen, dass sich diese Merkmale im Verlauf geologischer Zeit nicht viel verändert haben. Es gibt bis jetzt keinen Beweis, der die Richtigkeit dieser Behauptung widerlegt (oder sie auch nur ernsthaft bezweifelt)... Mehrere Beweislinien deuten darauf hin, dass es unwahrscheinlich ist, dass sich die Biochemie oder die Physiologie der Cyanobakterien seit mindestens ~2,1 Milliarden Jahren sehr verändert hat. Bis zum Beginn der recht gut beurkundeten fossilen Funde. Das fügt die cyanobakteriellen Fossilien in ein evolutionäres Kontinuum ein, das bis in die Gegenwart reicht." Schopf und Klein (1992:596, 597).
Hat sich dieses "evolutionäre Kontinuum", die erste lebende Zelle, dann in höhere Formen des Lebens entwickelt, in Pflanzen, Tiere und Menschen?
J. W. Schopf und C. Klein: "In bezug auf die Biochemie zum Beispiel: Die Spannweite der Formen in den Scheiden der Proterozoischen Cyanobakterien (von dünn, zerstreut, und transparent bis dick, robust, und viellamellig) gleicht im wesentlichen dem, was für die Scheiden kennzeichnend ist, welche die kugelförmigen und faserigen Cyanobakterien umschließen. Dann muss auch die ursprüngliche Polysaccharid-Zusammensetzung mit der der heutigen Zellen übereinstimmen. Wie man allgemein berichtet, haben sich besonders diese Scheiden in den proterozoischen fossilen Funden und in den tieferen Teilen heutiger mikrobieller Matten erhalten.
"Was die Physiologie betrifft: Das Vorkommen von isoprenoidem Kohlenwasserstoff Pristan und Phytan (zum Teil, vermutlich durch Chlorophyll entstanden) in fossilen stromatolitischen, proterozoischen Sedimenten, die Cyanobakterien enthalten, die Kohlenstoff-isotopischen Signaturen von Kerogen, die man aus diesen Einheiten isoliert hat, und die blätterige, Matten formende Orientierung stromatolitischer cyanobakterieller Photosynthese, der Sauerstoff, der so freigesetzt wurde. Das beweisen vermutlich die oxidierten proterozoischen Sedimente, wie die gestreiften Eisenformationen und roten Betten (Rotliegendes, rote Sandsteinschichten).
"Auch die Paläoökologie/Paleobathymetrie zeigt uns: Alle proterozoischen Sedimente, die nicht-transportierte fossile Cyanobakterien enthalten, scheinen in der photischen Zone (= wo es hell ist) abgesetzt worden zu sein. Die Fossilien gehören dort, wie die heutigen Cyanobakterien, einer stromatolitischen Lebensgemeinschaft an. Auf der Ebene der Familie oder der Art ist die Zusammensetzung der proterozoischen und heutigen Matten bauenden cyanobakteriellen Gesellschaften sehr ähnlich. Beide werden von Oscillaltoriazäen (Cyanobakterien) beherrscht (z.B., Oscillatoria, Lyngbya, Phormidium, Microcoleus). Untergeordnet sind dort die Chroococcazäen (z.B., Microsystis, Aphanocapsa, Gloeocapsa, Chroococcus) und, in einigen Fällen, Entophysalidazäen (z.B., Pleurocapsa).
"Schließlich, der spärliche Beweis, den wir jetzt in den Extrakten der organischen Geochemie haben (z.B., die proterozoische Verteilung von Hopanen, Steranen und anderen Lebenskennzeichen)... Das ist zwar nicht eindeutig. Doch das stimmt mit der Annahme überein, dass sich die Cyanobakterien biochemisch und physiologisch nicht sehr verändert haben.
"Indirekte Beweise und 'Übereinstimmungsargumente' wie die Vorangehenden sind zwar nicht zwingend. Doch es scheint vernünftig, dies anzunehmen: Die hypobradyletische (= wenig oder gar keine) Evolution der proterozoischen Cyanobakterien war nicht nur für ihre Morphologie kennzeichnend, sondern mindestens auch für die grundlegenden Aspekte ihrer Genetik, Biochemie und Physiologie." Schopf und Klein (1992:596, 597).
Cyanobakterien: seit wann?
Seit wann gibt es Cyanobakterien auf der Erde, die man früher als blau-grüne Algen bezeichnet hat? Wo haben sie gelebt? Und wie komplex waren sie?
J. William Schopf, am Molekularen Institut, Universität von Kalifornien, Los Angeles, berichtet in Science, 30. April 1993, S. 640-646 über seine neuen Forschungsergebnisse in Nordwest Australien:
"Elf Ordnungen (einschließlich acht bis jetzt unbeschriebene Arten) als Zellen erhaltene faserige Mikroben, die zu den ältesten Fossilien gehören, hat man in geschichtetem Kieselschiefer des Frühen Archaischen Apex-Basalts im nordwestlichen West Australien entdeckt. Dieser prokaryotische Fund beweist, dass schon damals trichomische cyanobacterium-ähnliche Mikroorganismen gelebt haben. Und dass sie schon so früh, vor mindestens ~3.465 Millionen Jahren, morphologisch verschiedenartig waren. Das deutet darauf hin, dass die Sauerstoff erzeugende Photoautotrophie schon so früh in der Geschichte des Lebens entstanden sein muss.
"Das vielversprechendste Gebiet für solche Studien ist der Pilbara Block im nordwestlichen West Australien. Dieses Gebiet liegt auf einer 30 km mächtigen Schichtenfolge von recht gut erhaltenen sedimentärem und vulkanischem Gestein. Sie sind ~3.000 bis 3.500 Millionen Jahre alt. Aus diesem Gebiet beschreibe ich eine verschiedenartige Ansammlung faseriger mikrobieller Fossilien. Man hat sie im Frühen Archaischen (~3.465 Millionen Jahre alten) Apex-Kieselschiefer entdeckt. Diese zellularen Prokaryoten sind mehr als 1.300 Millionen Jahre älter, als irgendein vergleichbarer Fund von Fossilien, den man bis jetzt aus den geologischen Berichten kennt.
"Cyanobakterielle, Sauerstoff erzeugende Photosynthetisierer mögen schon so früh auf der Erde gelebt haben. ... In einer Scheide steckende koloniale Einzeller findet man im ~3.465 Millionen Jahre alten sedimentären Gestein der Towers-Formation, auch in West Australien; und schmale, nicht durch eine Scheidewand getrennte bakterium-ähnliche Fäden von den ~3.450 Millionen Jahre alten Sedimenten der Swasiland Supergruppe von Südafrika.
"Ein maximales Alter für den Apex-Kieselschiefer von ~3.470 Million Jahren beweist ihr U-Pb Zirkon Alter... Ein Mindestalter für das fossilhaltige Gestein von ~3.460 Millionen Jahren zeigt uns das U-Pb-Zirkon Datum von 3.458 ± 1.9 Millionen Jahren für die Panorama-Formation. Sie liegt direkt darüber. ... Deshalb ist dieser fossilhaltige Apex-Kieselschiefer offensichtlich etwa 3.465 Millionen Jahre alt." (Schopf, J. W. 1993:640, 641).
Wie kann man wissen, ob dies wirklich die Überreste bakterieller Zellen sind? Wie kann man das wissenschaftlich beweisen?
Prof. J. W. Schopf: "Ihre offensichtlich zellulare Organisation und ihre morphologische Kompliziertheit und Ähnlichkeit mit jüngeren Prokaryoten, mit fossilen und modernen, beweisen klar, dass sie von Lebewesen stammen. ... Ob die Mikrofossilien sehr viel älter sind, oder ob sie ebenso alt sind wie der Apex-Kieselschiefer, weiß man nicht." (1993:641, 642).
"Elf Ordnungen der faserigen, dunkelbraun bis schwarzen kohlenhaltigen Mikrofossilien, einschließlich acht neue Arten, hat man in dieser Ablagerung identifiziert. Einzeln lebende einzeller-ähnliche Gebilde, von kugeliger Form, aber unsicheren biologischen Ursprunges, kommen auch vor.
"Einzelne Ordnungen (besonders, Primaevifilum minutum, n. sp.; P. laticellulosum, n. sp. und P attenuatum n. sp.) oder besondere Paare oder Gruppen von Ordnungen (zum Beispiel, P. delicatulum und Archaeoscillatoriopsis disciformis, n. gen. n. sp.; oder P. delicatulum, P. amoenum und P. conicoterminatum) sind gewöhnlich in den einzelnen Trümmergesteinen am zahlreichsten. Sie gehören wahrscheinlich zu einer benthischen (= am Meeresboden lebenden) mikrobiellen Population. Sie war nur lose organisiert und war in Schleim eingebettet. Die Fäden zeigen dort weder die subparallele Orientierung noch die laminare Organisation, die für die meisten stromatolitischen Mikrobiotope kennzeichnend ist. Mikrofossilien hat man in dem stromatoliten-ähnlichen, geschichteten Trümmergestein nicht gefunden, die dort auch vorkommen. ... Die unvollständige Bewahrung der Apex-Fossilien zeigt uns, dass die ursprüngliche Ansammlung wahrscheinlich mehr Ordnungen umfasst hat, als die 11 Arten, die man bestimmt hat.
"Wir können heute beweisen: Die fossilen Aufzeichnungen sind mehr oder weniger ununterbrochen und recht gut bekannt. Und zwar von etwa 2.100 Millionen Jahren an bis heute. Sie beginnen mit den verschiedenartigen Mikrobiotopen der ~2.100 Millionen Jahre alten Belcher Gruppe und der ~2.080 Millionen Jahre alten Gunflint Eisenformation, beide in Kanada. Doch die fossile Aufzeichnung von den mehr als 1.300 Millionen Jahren, die zwischen diesen Ablagerungen und dem Apex-Kieselschiefer liegt, ist praktisch nicht entziffert. Es gibt zwar diese große Lücke in der Aufzeichnung. Aber die morphologische Ähnlichkeit der Apex-Fossilien mit den geteilten, faserige Prokaryoten, proterozoisch und modern, zeigt uns, dass sie fast bestimmt Prokaryoten sind. Sie gehören einer evolutionären Zeitspanne an, das sich vom Frühen Archaikum bis zur Gegenwart erstreckt. Diese Ansicht ist dadurch erhärtet, weil in den Apex-Fäden Zellen vorkommen, die sich teilen und Zellpaare. Dabei handelt es sich offensichtlich um teilweise Teilung, also um Zellteilung, wie das auch heute in prokaryotischen Fäden geschieht." (1993:642, 643).
Wie unterscheiden sich diese, etwa 3,5 Milliarden Jahre alten, bakteriellen Zellen, von den heutigen bakteriellen Zellen?
Prof. J. W. Schopf: "Im Vergleich zu modernen Prokaryoten ähneln die meisten Apex-Mikroben besonders trichomischen (nicht-umhüllten oder dünn umhüllten) Oscillatoriazäen-Cyanobakterien. Die Zellweiten der Apex-Ordnungen reichen von 0,5 µm bis zu 19,5 µm und sind durchschnittlich ~5,0 µm breit (Tabelle 1). Moderne faserige Bakterien sind gewöhnlich ganz schmal, vorwiegend < 1,5 µm im Durchmesser. Die Trichome (= Fäden) der meisten Oscillatoriazäen sind viel breiter. Aufgrund von morphometrischen Analysen von mehr als 500 Arten moderner faseriger Mikroben habe ich vorgeschlagen, dass man fossile septierte Fäden < 1,5 µm breit als 'wahrscheinliche Bakterien' betrachtet. Jene, die 1,5 µm bis 3,5 µm breit sind, als (undifferenzierte) 'Prokaryoten.' Und jene, die > 3,5 µm breit sind, als 'wahrscheinliche Cyanobakterien'.
"Wendet man diese Kriterien auf die Apex-Fossilien an, dann bedeutet das für mich: Zwei Arten (Archaeotrichion septatum, n. sp., und Eoleptonema apex, n. sp.) sind wahrscheinlich Bakterien; zwei Arten (Primaeevifilum minutum, n. sp., und P. delicatum) sind entweder Bakterien oder Cyanobakterien; und die übrigen sieben Arten, fast zwei Drittel der Ordnungen (und 63 Prozent der gemessener Exemplare), sind wahrscheinlich Cyanobakterien.
"Die Größenspanne der faserigen Bakterien und Cyanobakterien überschneidet sich. Deshalb ist diese Einteilung nicht absolut. Das Muster der Größenverteilung in der Apex-Ansammlung ähnelt mehr den heutigen Oscillatoriazäen, als den nicht-cyanobakteriellen-Prokaryoten. Mehrere Apex-Arten, insbesondere jene mit breiten Trichomen (Primaevifilum laticellulosum, n. sp.; Archaeoscillatoriopsis grandis, n. gen., n. sp.; und A. maxima, n. gen. n. sp.) unterscheiden sich in ihrer Zellgröße von fast allen Bakterien. Doch man kann sie praktisch nicht von bestimmten Oscillatoriazäen unterscheiden, von Proterozoischen (Oscillatoriopsis spp.) und modernen (Oscillatorioa spp.). Wenn man die Apex-Fäden in späteren Präkambrischen Sedimenten entdeckt hätte, in denen fossile Oscillatoriazäen gut bekannt und recht weit verbreitet sind, oder wenn man sie in einer modernen mikrobiellen Gesellschaft entdeckt hätte, und wenn ihre Morphologie das einzige Kriterium wäre, durch das man auf ihre biologische Verwandtschaft schließen könnte, dann würde man die meisten als Oscillatoriazäen-Cyanobakterien betrachten." Schopf, J. W. (1993:643).
"Die Spannbreite der Morphologien der Apex-Fäden zeigt uns: Wenn die meisten Oscillatoriazäen sind, dann kann diese primitive Familie faseriger Cyanobakterien vorübergehend auf anoxische (baterielle) Photosynthese umschalten. Die Sauerstoff erzeugende Photoautotrophie ist vermutlich ein universales, sich selbst entwickelndes Merkmal dieser Gruppe. Die Gegenwart der verschiedenartigen Oscillatoriazäen in der Apex-Ansammlung deutet wahrscheinlich an, dass dieses recht fortschrittliche Niveau physiologischer Evolution schon vor mindestens ~3.465 Millionen Jahren erreicht worden ist." (1993:643, 644).
Was beweist, dass Sauerstoff erzeugende Oscillatoriazäen (= Cyanobakterien) schon so früh auf der Erde, vor etwa 3.500 Millionen Jahren, gelebt haben?
Prof. J. W. Schopf: "Vier andere Beweislinien scheinen die Existenz von O2 erzeugenden Oscillatoriazäen im Frühen Archaikum zu bestätigen: (i) Früh Archaische Stromatoliten wurden vermutlich von Photoautotrophen dominierte mikrobielle Gesellschaften produziert. (ii) Die Reaktionsmittel, die für die oxygene Photosynthese nötig sind, CO2 und H2O, und Materialien, die Produkte, die daraus stammen, sedimentäre organische Materie und oxydierte Eisen-Mineralien, waren in der Frühen Archaischen Umwelt vorhanden. (iii) Die isotopische Zusammensetzung der Frühen Archaischen organischen und kohlensauren Kohlenstoffes deutet darauf hin, dass er von der photosynthetischen CO2-Fixierung stammt. Auch in heutigen mikrobiellen Populationen gibt es recht hohe CO2-Konzentrationen. (iv) Kalkulationen, die auf Modellen des frühen globalen Ökosystems beruhen, und auf der Cerium und Europium-Konzentration in Archaischen gestreiften Eisenformationen, deuten darauf hin, dass die O2 erzeugende Photosynthese und die aerobe Atmung, beide aus dem Frühen Archaikum stammen. Diese zusätzlichen Beweislinien sind aber nicht überzeugend. Alle, außer der letzten, die notwendigerweise Modell-abhängige Ungewissheiten enthält, könnten auch von bakteriellen Photosynthetisierern stammen." (1993:644).
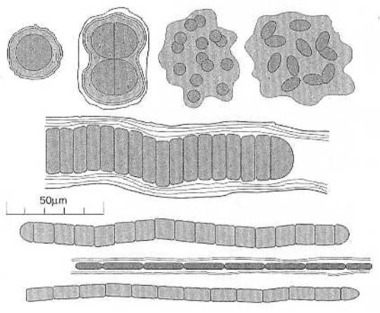
Cyanobakterien, coccoidal (kugelförmig), ellipsoidal und fadenförmig. Cyanobakterien gibt es in vielen Formen und Größen. Oft sind sie von einer schleimigen Schicht, die sie ausgeschieden haben, umhüllt, oder sind darin eingebettet. Jede dieser Scheiben in diesen Faden oder Wurst ähnlichen Gebilden ist ein fotosynthetisches Cyanobakterium. Das Cyanobakterium verwandelt die Energie des Sonnenlichts in organische Stoffe. Und es erzeugt freien Sauerstoff. Nach J. W. Schopf (1999:197) Bild 7.1. Das erste Cyanobakterium, vor etwa 3,7 Milliarden Jahren, war schon ebenso komplex wie das heute lebende. Es hat sich überhaupt nicht in irgendwelche höhere Formen des Lebens entwickelt, weil es schon damals seiner Umwelt vollkommen angepasst war.
Cyanobakterium
Auch die versteinerten Überreste von verschiedenen Arten des Cyanobakteriums, des grünlich-blauen Bakteriums, hat man in Nordwest Australien gefunden. Das Cyanobacterium Oscillatoria, hat man bei Warrawoona in N.W. Australien gefunden. Es ist etwa 3,3-3,5 Milliarden Jahre alt. Professor J. W. Schopf (1983, 1992) hat über diese Funde berichtet. Wie groß sind diese Sauerstoff erzeugenden Cyanobakterien? Wie lang ist ihr Genom, ihr genetischer Code? Und wie viel Information enthält es? Mit anderen Worten: Was war nötig, um ihre DNA-Paare richtig zu ordnen? Betrachten wir kurz einigen von ihnen:
Primaefilum conicoterminatum, 4,5 bis 7,0 µm breit und 3,0 bis 4,5 µm lang. Durchschnittlich 5,9 µm breit und 3,9 µm lang. Schopf und Klein (1992:35). Ihm entspricht das heute lebende Cyanobakterium Oscillatoria acuta.
Primaevifilum laticellulosum, 6,0 bis 8,5 µm breit und 2,5 bis 50 µm lang, durchschnittlich 7,0 µm lang und 3,5 µm breit. Von Warrawoona, Apex-Kieselschiefer, 3,5 Milliarden Jahre alt. Ihm entspricht das heute lebende Cyanobakterium Oscillatoria tenus. Schopf J. W. (1993:641, 645).
Gattung Archaeoscillatoriopsis, 3,0 bis 19,5 µm breit 0,8 bis 6,0 µm lang. Die entsprechende heutige Form: Oscillatori ssp. oscillatoriaceans. Warrawoona, Apex-Kieselschiefer, 3,5 Milliarden Jahre alt. Schopf, J. W. (1993:641, 645).
Archaeoscillatoriopsis disciformis, 3,0 bis 5,5 µm breit, 0,8 bis 2,2 µm lang. Durchschnittlich 4,2 µm lang, 1,2 µm breit. Entsprechende heutige Form: Cyanobakterium Oscillatoria grunowiana. Von Warrawoona, Apex-Kieselschiefer, 3,5 Milliarden Jahre alt. Schopf, J. W. (1993:641, 645).
Archaeoscillatoriposis grandis, 8,0 bis 11,5 µm breit, und 1,0 bis 3,5 µm breit. Durchschnittlich 9,0 µm lang, 2,0 µm breit. Entsprechende heutige Form: modernes Cyanobakterium Oscillatoria chalybea. Schopf, J. W. (1993:641, 645). Von Warrawoona, Apex-Kieselschiefer, 3,5 Milliarden Jahre alt.
Archaeosillatoriopsis maxima, 15,0 bis 19,5 µm breit und 3,0 bis 6,0 µm lang. Durchschnittlich 16,5 µm breit und 4,5 µm lang. Entsprechende heutige Form: modernes Cyanobakterium Oscillatoria antillarum. Von Warrawoona, Apex-Kieselschiefer, 3,5 Milliarden Jahre alt. Schopf, J. W. (1993:641, 645).
Wie lang ist das Genom dieses Sauerstoff erzeugenden Cyanobakteriums? Und wie viel Information enthält es? - Michael Herdman und Mitarbeiter (1979:80) haben die Genomgröße von sechs Rassen von Oscillatoria bestimmt. Sie schwankt zwischen 2,50·109 bis 4,38·109 Dalton. Seine durchschnittliche Größe ist 3,62·109 Dalton. 3,62·109 Da : 660 Da/bp = 5.484.848 bp log 4 = 103.303.207 Bit.
Das bedeutet: Schon vor 3,5 Milliarden Jahren hatte sich die Erde genug abgekühlt. Es gab viele verschiedene Gattungen und Rassen des Cyanobakteriums Oscillatoria. Viele dieser Arten leben noch heute. Sie haben sich überhaupt nicht verändert, weil sie von Anfang an vollkommen ihrer Umgebung angepasst waren! Ihr Genom hatte eine durchschnittliche Größe von 5.484.848 Basenpaaren. Die meiste genetische Informationen der Zelle ist in ihrem Genom gespeichert. Es enthält mindestens 103.303.207 Bit Information oder ja/nein Entscheidungen!
Synechocystis
Auch das Sauerstoff erzeugende Cyanobakterium Synochocystis aquatilis hat man in den 3,3-3,5 Milliarden Jahre alten Sedimenten bei Warrawoona, in N.W. Australien gefunden. Es hat einen Durchmesser von 5,0 bis 6,0 µm. Schopf, J. W. (1983) Tabelle 9.2. Wie lang ist sein Genom? Und wie viel Information enthält es?
Synechocystis, mit einem Durchmesser von 6-7 µm (PCC 6806) hat eine Genomgröße von 2,31·109 Da. Eine Zelle mit einem Durchmesser von 5,0-6,0 µm hat dann eine Genomgröße von etwa 2·109 Da. Herdman, M. et al. (1979:67). 2·109 Da : 660 Da/bp = 3.030.303 bp log 4 = 101.824.424 Bit.
Synechocystis hat jetzt eine Genomgröße von 1,79·109 bis 3,5·109 Da. Seine mittlere Größe ist 2,29·109 Da. 2,29·109 Da : 660 Da/bp = 2.712.121 bp log 4 = 102.088.965 Bit.
Ergebnis
Warum gibt es Leben auf der Erde? Warum gibt es einzellige Organismen? Was zeigen uns die Funde von der Warrawoona Gruppe im nordwestlichen Australien, die 3,3-3,5 Milliarden Jahre alt sind? Beweisen sie, dass sich das Leben auf der Erde aus einem gemeinsamen Vorfahren, aus der ersten Zelle, entwickelt hat?
Die Fossilen von Warrawoona widerlegen eindeutig den Glauben der Evolutionisten. Es gibt überhaupt keinen Beweis dafür in den Sedimenten der Frühen Präkambrium-Zeit vor etwa 3,3-3,5 Milliarden Jahren, dass sich alles Leben auf der Erde aus einem gemeinsamen Vorfahren, aus der "ersten Zelle" entwickelt hat. Im Gegenteil. Schon von Anfang an gibt es viele verschiedene Gattungen und Arten von Bakterien und Archaebakterien, wie heute.
Es gibt überhaupt keinen Beweis in den ältesten Sedimenten der Welt, dass sich anorganische Materie aus Nicht-Lebendem in Lebendes verwandelt hat, vom Niedrigen zum Höheren, und vom Einfachen zum Komplexen. Im Gegenteil. Nur Überreste perfekter einzelliger Organismen, von Bakterien und Archaebakterien hat man gefunden. Sie sind mit vielen der heute lebenden Gattungen und Arten identisch. Sie wissen nichts von Evolution. Sie sind geblieben, was sie von Anfang an gewesen sind. Auch die gestreiften Eisenformationen (BIFs) und der organische Kohlenstoff von Warrawoona beweisen: Diese ersten Zellen waren schon ebenso komplex, wie die Zellen, die jetzt, etwa 3,3-3,8 Milliarden Jahre später, leben.
In den Sedimenten des Frühen Präkambriums gibt es keinen Übergang vom Nicht-Lebenden zum Lebenden, vom einfachsten Atom und den Molekülen zum Menschen. Es gibt nur perfekte einzellige Organismen, die ihrer Umgebung vom Anfang vollkommen angepasst waren. Die erste lebende Zelle auf Erde war schon komplexer, als irgend etwas, was der Mensch bis jetzt geschaffen hat. Die Funde aus der Zeit, als das Leben auf der Erde erschien, beweisen eindeutig, dass Gott sie erdacht und erschaffen hat. Was haben andere Wissenschaftler über das Leben auf der Erde herausgefunden. Und wie ist es entstanden?
Proterozoische und lebende Cyanobakterien
Cyanobakterien aus der Frühen Proterozoischen Zeit, vor etwa 2.000 bis 2.500 Millionen Jahren, nach der Archaischen Zeit: Wie komplex waren diese einzelligen Organismen? Wie sehr haben sie sich seitdem verändert? Haben sie sich vielleicht in höhere Formen des Lebens entwickelt?
Andrew H. Knoll, Botanisches Museum, Harvard Universität, Cambridge, MA, USA, und Stejepko Golubic, Abt. Biologie, Boston Universität, Boston, MA, USA, berichten über "Proterozoische und lebende Cyanobakterien":
"Im Grunde genommen findet man alle wichtigen morphologischen Merkmale, die man bei der taxonomischen Einteilung lebender Cyanobakterien gebraucht, auch in wohlerhaltenen Mikrofossilien. Trotz des großen Einflusses der taphonomischen Änderungen proterozoischer mikrobieller Überreste. (Taphonomisch: als sie starben und begraben wurden). ... Einige Blau-Grüne (= Cyanobakterien) bilden extrazellulare Scheiden oder Hüllen, die gegenüber Änderungen nach dem Tode recht immun sind. Diese Überreste können sich drei-dimensional erhalten. Dadurch lernen wir ausführlich über Zellgröße, Form, und Faden von koloniellen Formen." (1992:453).
Eoentophysalis
A. H. Knoll und S. Golubic: "Einer der ersten, und immer noch einer der aufschlußreichsten Vergleiche einer proterozoischen Mikrofossil-Population und einem bestimmten heutigen Cyanobakterium ist der zwischen der fossilen Art Eoentophysalis und seinem modernen Gegenstück, Entophysalis. ... Die Gattung Entophysalis wird durch seine kugelförmigen Zellen charakterisiert, die durch binäre Spaltung in drei Ebenen entstehen. Äußere Hüllen sind bei mehreren Zellteilungen vorhanden. Daraus entstehen dann Zellpakete. Sie bewahren ihre Teilungsgeschichte.
"Moderne Entophysalis Arten gibt es in verschiedenen peritidalen (= Gezeiten) und Playa-See-Umgebungen. Am Rand von Gewässern, die sehr salzig sind, sind sie besonders zahlreich und auffallend. In solchen Umgebungen können Entophysalis Bevölkerungen strukturell fähige und sedimentologisch auffällige Matten bauen. Und sie können sich am Aufbau und der Versteinerung stromatolitischer Strukturen beteiligen." (1992:455).
"Entophysalis ist ein Cyanobakterium, das morphologisch, ökologisch und taphonomisch gut charakterisiert ist. Wichtig ist auch. Es ist ein Cyanobakterium, dessen Morphologie für die Ordnung kennzeichnend ist, (d.h., die auffällige Kolonie-Form von Entophysalis findet man in keinem anderen eubakteriellen Stamm). Man kann es deshalb gut mit fossilen Populationen vergleichen. Forschungsberichte, die man in den letzten 15 Jahren veröffentlicht hat, zeigen, dass nahe fossile Gegenstücke zu Entophysalis in proterozoischen Schichten weit verbreitet sind.
"Die Gattung, Eoentophysalis hat man für große Populationen in der Frühen proterozoischen Belcher Supergruppe in Kanada aufgestellt (Hofmann 1976). Die Belcher Eoentophysalis Populationen stimmen mit der lebenden Entophysalis Art in ihrer Zellgröße und -form, Teilungsmuster und Lebenszyklus, Art der Hülle und Kolonieform, taphonomischem Muster und Ökologie und Umweltverteilung überein (Golobic und Hofmann 1976). Sie lassen wenig Zweifel an der systematischen Paläontologie und den physiologische Eigenschaften der beinahe 2.000 Millionen Jahre alten Fossilien. Eoentophysalis Populationen, die mehrere Arten umfassen, hat man auch aus dem Mittel proterzoischen McArthur beschrieben (Muir 1976), Nathan (Oehler 1978), und Dismal Lake Gruppen (Horodysk und Donaldson 1983), und Gaoyuzhuan Formation (Zhang 1981). Späte proterozoische Beispiele kennt man von der Narssarssuk (Strother et al. 1983) und Bitter Springs Formation (Knoll und Golubic 1979). ... In allen Fällen entsprechen die fossilen Bevölkerungen morphologisch, ökologisch und taphonomisch ihrem lebenden Gegenstück." Knoll, A. H. und S. Golubic (1992:455).
Polybessurus
Knoll und Golubic: "Morphologisch auffällige Mikrofossilien, die man zur Gattung Polybessurus gestellt hat, hat zuerst Fairchild von den verkieselten Ober-Proterozoischen karbonisierten Salzen von Süd Australien beschrieben (1975). In jüngster Zeit haben Green et al. (1978) ausführliche Beobachtungen und Beurteilungen großer Populationen von der Ober-Proterozoischen Eleonore Bucht Gruppe, Ost Grönland, veröffentlicht. Wie im Fall von Eoentophysalis, passen auch die Polybessurus Populationen mit lebendem Cyanobakterien zusammen. Deren Morphologie ist für diese Ordnung kennzeichnend. (Green et al. 1987).
"Polybessurus Individuen waren kugelförmige Einzeller. Deren systematische Hüllenproduktion bildete einen gallertartigem Stiel, der die Zelle über die Sedimentoberfläche hob. Fortgepflanzt haben sie sich durch Baeozyt-Formation. Polybesssurus Fossilien kommen manchmal als isolierte Individuen innerhalb mikrobieller Matten vor. Doch auffälliger war: Große Populationen bildeten auf feinkörnigen, kohlensauren Oberflächen in peritiden (Gezeiten) Umgebungen Krusten. Heute kennt man Populationen, die zu dieser Beschreibung passen, aus Mittel und Ober Proterozoischen Formationen in Ost Grönland (Green et al. 1987), Svalbard (Butterfly und Knoll, unpubl. Daten), Australien (Fairchild 1975), und dem Arktischen Kanada (Butterfield et al., unpubl. Daten).
"Physisch ähnliche Umgebungen auf der heutigen Bahama Bank beherbergen Cyanobakterien. Sie sind etwas kleiner, und entsprechen den Proterozoischen Polybessurus Populationen (Green et al. 1987). Dieses moderne Gegenstück hat man noch nicht beschrieben. Aber es ähnelt der Gattung Cyanostylon. Es bildet gallertartige Stiele durch asymmetrische Gel-Ausscheidung, wenn nacheinander auf ihrer oberen, dünneren Seite die Hüllen aufbrechen. Es entsteht dann ein Stapel umgekehrter gallertartiger Kegel. Das Vorkommen dieser lebenden Cyanobakterien in Umgebungen, die man auf der Basis Proterozoischen fossilen Aufzeichnung vorhergesagt hat, lässt auch hier wenig Zweifel an der systematischen und ökologischen Auslegung der Fossilien." Knoll und Golubic (1992:455, 456).
Eohyella
Knoll und Golubic: "Wir möchten hier kurz einen dritten wichtigen alten/modernen Vergleich anführen, um diesen Punkt zu befestigen. Ooide, die man heute in seichten subtidalen Umgebungen der Bahama Bank gefunden hat, sind mit endolithischen Cyanobakterien übersät; bis zu sechs Arten kann man in einem einzigen Loch (Coid) finden. Diese Cyanobakterien sind morphologisch und von ihrer Entwicklung und ihrem Verhalten her auffällig. Spät-Proterozoische Ooiden von der Eleonore Bucht Gruppe, Ost Grönland (Green et al. 1988), und der Backlundtoppen Formation, Svalbard (Knoll et al. 1989), beherbergen viele und verschiedenartige endolithische Mikrofossilien. Ihre Morphologie, Entwicklung und Fortpflanzung, Orientierung innerhalb des Substrates (von dem man auf ihr Verhalten schließen kann), und ihre Verbreitung in der Umwelt, entsprechen genau denen von Populationen, die man in heutigen Ooiden gefunden hat.
"Es gibt mehrere Arten der cyanobakteriellen Gattung Hyella in Bahamischen endolithischen Gesellschaften. Diese Populationen sind durch mehrere Merkmale miteinander verbunden: das Vorhandensein von Pseudofäden, die aus kugelförmigen bis zu zylindrischen kugelförmigen Zellen, Pseudo-Faden-Verzweigung durch Zellteilung oder apikale Teilung der Zelle in zwei Zellen, Hüllen, die nicht zerfallen. Sie bestimmen die Formen und Positionen der Zelle. Binäre Zellteilung, die von Baeozyt-Formation begleitet wird, und ein auffälliger negativer Phototropismus und eine Polaritätsumkehr in der Entwicklung.
"Vier Arten der Gattung Eohyella kommen in Spät-Proterozoischen Ooiden von Svalbard und Ost Grönland vor. Zwei prominente Populationen sind eng mit Hyella gigas und H. caespitosa vergleichbar. Und ein drittes lebendes Gegenstück gehört einer Hyella Art an, die man noch nicht beschrieben hat. Dieser Vergleich über 700-800 Millionen Jahren hinweg entspricht dem oben angeführten. Doch hier kommen zum Verhalten noch Morphologie, Lebensstil, und Umgebung in der Beurteilung. Eohyella kommt in der Tat in 1.700 Millionen Jahren alten Stromatoliten in China vor (Zhang und Golubic 1987). Aber bestimmte Vergleiche mit heutigen Arten kann man noch nicht machen." Knoll und Golubic (1992:456, 457).
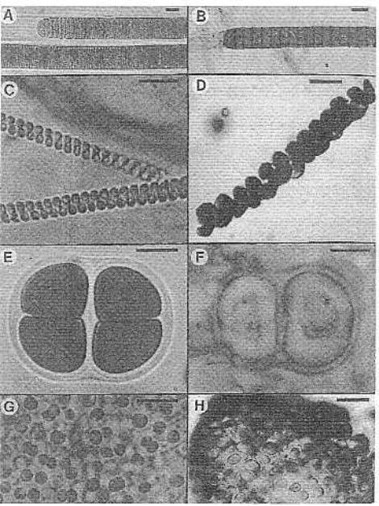
Sie haben sich seit Milliarden von Jahren nicht verändert. Lebende Cyanobakterien, die Stromatoliten bauen, aus Nordmexiko (A, C. E, und G) und ihre präkambrischen Ebenbilder (look-alikes) (B, aus der 950-Jahre-alten Lakhanda Formation, und D, der 850 Millionen-Jahre-alten Miroedikha Formation, beide aus Siberien; F, der 1,550-Millionen-Jahre-alten Satka Formation in Baschkirien; and G, der 2,100-Millionen-Jahre-alten Belcher Supergruppe in Kanada). (A) Lynbya, verglichen mit (B) Palaeolynbya. (C) Spirulina, verglichen mit (D) Heliconema. (E) Gloeocapsa, verglichen mit (F) Gloeodiniopsis. (G) Entophsalis, verglichen mit (H) Eoentophysalis. Maßstab = 10 µm. Aus: J. W. Schopf, Cradle of Life (1999) Bild 8.10.
Alte fossile Zellen: ihre DNA
Wie komplex waren die alten einzelligen Organismen, deren fossile Überreste man jetzt in verschiedenen Teilen der Welt gefunden hat? Wie viel Information haben die Bakterien und Archaebakterien enthalten, die lange vor uns, vor Milliarden von Jahren, auf unserem Planeten Erde gelebt haben? Wie lang war ihr Genom (DNA-Strang)? Und viel Information braucht man, um seine Nukleotide an die richtige Stelle zu setzen? In anderen Worten: Was sind die Sequenz-Alternativen für ihr Genom, so dass jedes Basenpaar an die richtige Stelle kommt?
Beggiatoa
Bei Warrawoona, West Australien, hat man auch die fossilen Überreste des Bakteriums Beggiatoa arachnoidea und Beggiatoa minima gefunden. Sie haben dort vor etwa 3,3-3,5 Milliarden Jahren gelebt (Schopf, J. W. 1983, Tabelle 9.2). Der größte Teil der genetischen Information der Zelle ist in ihrem Genom, auf ihrem DNA-Strang, verschlüsselt. Wie lang war ihr Genom? Und wie viel Information braucht man, um seine Nukleotid-Paare richtig zu ordnen?
Wir können das jetzt nur auf der Ebene der Gattung herausfinden. Weil man bis jetzt nur die Genomgröße von Beggiatoa alba veröffentlicht hat. Es hat 2,02·109 Da = 3,03·103 Kilobasenpaare, gemäß Genthner et al. 1985, Bergey's Manual (1989:2092).
2,02·109 Da : 660 Da/bp = 3.060.606 bp
3.060.606 log 4 = 101.842.668 Bit Information, Reihenfolge-Alternativen.
Das bedeutet: Mindestens 101.842.668 Bit Information (ja/nein Entscheidungen) braucht man, um die Basenpaare im DNA-Strang der Gattung Beggiatoa richtig zu ordnen. Wir sollten hier bedenken: Alles Wissen, das der Mensch bis jetzt in Büchern aufgeschrieben hat, umfasst "nur" 1018 Bit (Gitt, W. 1986:68). Die Unterschiede innerhalb einer Gattung einer bakteriellen Zelle sind nur rassische Variationen ihrer Art. Das hat nichts damit zu tun, dass sie sich in höhere Arten von Pflanzen und Tieren entwickelt hat.
Methanococcus
Bei Warrawoona, West Australien, hat man auch die fossilen Überreste der Archaebakterie Methanococcus vaniellii gefunden. Sie hat dort vor etwa 3,3-3,5 Milliarden Jahren gelebt (Schopf, J. W. 1983, Tabelle 9.2). Man hat die Genomgröße von zwei anderen Spezies von Methanococcus veröffentlicht, von M. thermolithoautotrophicum und M. voltae. Methanococcus thermolithoautotrophicum SN 1 hat eine Genomgröße von 1,1·109 Da, wie A. Klein und M. Schnorr (1984:630) berichten.
1,1·109 Da : 660 Da/bp = 1.666.666 bp log 4 = 101.132.044 Bit.
Das Genom von Methanococcus voltae hat 1.870.000 bis 1.899.000 Basenpaare, durchschnittlich 1.880.286 bp, wie J. Sitzman und A. Klein (1991:505-513) berichten.
1.880.286 bp log 4 = 101.132044 Bit.
Das bedeutet: Wenn wir den niedrigeren Wert von 101.003.432 Bit nehmen, dann brauchte man mindestens 101.003.432 Bit Information (oder ja/nein Entscheidungen), um den DNA-Strang der Archaebakterie Methanococcus zu machen. Soviel Information hat man gebraucht, nur um die Basenpaare seines Genom richtig zu ordnen, und zwar schon vor 3,2-3,5 Milliarden Jahren! Woher stammt diese Information?
Cyanobakterium
Die Bakterien haben sich überhaupt nicht in höhere Formen des Lebens entwickelt. Denn sie waren von Anfang an vollkommen ihrer Umwelt angepasst. Die Bakterien, deren versteinerte Überreste man in 3,3-3,5 Milliarden Jahren alten Sedimenten gefunden, kann man sogar nach ihrer Gattung bestimmen, und einige sogar nach ihrer Spezies. Das heißt, mit Gattungen und Spezies, die heute noch leben. Wenn sie sich durch Mutation und Selektion höher entwickelt hätten, wie uns das Evolutionisten gern einreden möchten, was sollten wir dann erwarten?
Sollten wir dann nicht erwarten, dass man für jede geeignete bakterielle Zelle, Hunderten oder Tausenden ungeeigneter Zellen findet, in all ihren verschiedenen Phasen der Entwicklung? Sollten wir dann nicht erwarten, dass die Sedimente unserer Erde hauptsächlich bakteriellen Abfall enthalten, wie eine riesige Müllkippe? Aber wir finden statt dessen nur perfekte Zellen seit dem Anfang, keinen biologischen Müll, noch irgendwelche dazwischen liegende Phasen. Wir finden auch keine Spur von einem primitiven "gemeinsamen Vorfahren alles Lebens". Den gibt es nur in der Phantasie leichtgläubiger Evolutionisten. Sie haben überhaupt keinen experimentellen oder beobachtbaren Beweis für ihre Behauptung. Das ist nur eine fromme Mythe.
Nicht nur die ältesten versteinerten Zellen, etwa 3,5 Milliarden Jahre alt, die wir jetzt kennen, sondern auch die Zellen, die man später in jüngeren Sedimenten gefunden hat, 2 Milliarden und 1 Milliarde Jahre alt, sind ebenso perfekt. Viele von ihnen hat man sogar nach Spezies bestimmt, die heute noch leben. Schauen wir uns jetzt kurz einigen Cyanobakterien an.
Lynbya
Prof. J. W. Schopf vergleicht die entdeckten Warrawoona Zellen mit denjenigen, die noch heute leben. Unter den vielen verschiedenen Gattungen und Spezies von Bakterien (und Archaebakterien), erwähnt er auch das Sauerstoff erzeugende Cyanobakterium Lynbya cryptovaginata. Es ist 3,3-3,5 Milliarden Jahre alt. Es hat einen Durchmesser von 4,0-9,0 µm. Bei Warrawoona hat man auch fünf verschiedene Spezies des Cyanobakteriums Phormidium gefunden. Sie haben einen Durchmesser von 0,6-6,7 µm, und eine Länge von 2,5-8,0 µm. Schopf, J. W. (1983) Tabelle 9.2. Man stellt sie jetzt mit Lyngbya in eine Gruppe. Mann N. H. et al. (1992:34).
In der Lakhanda Formation vom Khabarovsk Gebiet in Ost Sibirien, hat man Palaeolyngbya, ein Lyngbya-ähnliches fossiles Cyanobakterium gefunden. Es ist etwa 950 Millionen Jahre alt. Vergleichen Sie, bitte, Abb. 2.8A mit 2.8B in Schopf, J.W. (1992:51). Wie groß ist das Genom des Sauerstoff erzeugenden Cyanobakteriums Lyngbya? Wie viel Information enthält es?
Das Genom von Lyngbya (Rasse PCC 7419) ist etwa 4,58·109 Dalton groß, wie Bergey's Manual (1989:1777), und Herdman, M., et al. (1979b) berichten. 4,58·109 Da : 660 Da/bp = 6.939.393 bp log 4 = 104.177.930 Bit.
Spirulina
Das ist ein Sauerstoff erzeugendes Cyanobakterium. Die Überreste von Spirulina hat man in der 850 Millionen Jahre alten Miroedikha Formation im Turuchansk Gebiet von Ost Sibirien gefunden. Sehen Sie sich, bitte, die Bilder in Abb. 2.8C und 2.8D in J. W. Schopf (1992:51) an. Wie groß ist sein Genom? Das Genom von Spirulina hat 2,53•109 bis 5,19•109 Da, und durchschnittlich 3,86·109 Da. 3,86·109 Da : 660 Da/bp = 5.848.484 bp log 4 = 103.521.138 Bit. Genomgröße von M. Herdman et al. (1979:80). Man braucht hier 103.521.128 ja/nein Entscheidungen, um den DNA Strang dieser Zelle zu machen. Das heißt, seine Basenpaare richtig zu ordnen. Zufall, Mutation und Selektion können das nicht.
Gloeocapsa
Gloeodiniopsis ist ein Gloeocapsa-ähnliches fossiles Cyanobakterium. Es kommt aus der 1.550 Millionen Jahre alten Satka Formation der südlichen Ural Berge in Bashkirien. Schauen Sie sich, bitte, die Bilder alter und neuer Zellen in Schopf, J. W. (1992:51) an, Abb. 2.8E und 2.8F. Es hat sich überhaupt nicht entwickelt. Die Fossilien widerlegen eindeutig die Lehre der Evolution. Sie zeigen uns, dass das nur eine fromme Mythe ist. Wie groß ist das Genom von Gloeocapsa? Wie viel Information enthält es?
Das Genom von Gloeocapsa enthält 2,90·109 bis 3,47·109 Da, durchschnittlich 3,20·109 Da, wie M. Herdman et al. (1979:77) berichten. 3,20·109 Da : 660 Da/bp = 4.848.484 bp log 4 = 102.919.078 Bit.
Das bedeutet: 102.919.078 ja/nein Entscheidungen hat man gebraucht, um die Basenpaare seines Genoms richtig zu ordnen. Dies ist etwa so, als wenn man beim Schreiben die Buchstaben und Wörter an die richtige Stelle setzt. Wie viel Information ist das? Wir verstehen das etwas besser, wenn wir uns daran erinnern, wie viel Information der Mensch bis jetzt in Büchern aufgeschrieben hat. "Nur" 1018 Bit (W. Gitt, 1986). Der Zufall kann keine Information erzeugen. Aber er kann Information zerstören. Je mehr Fehler ein Computerprogramm enthält, um so unbrauchbarer wird es. Auch tausend Affen, die auf ihren Schreibmaschinen ein Shakespeare-Gedicht schreiben, erzeugen keine Information. Wichtig ist hier nur der Produzent (und Sender) der Information, nicht der Empfänger (M. Himmelheber).
Ergebnis
Von der Zeit an, wo unser Planet Erde entstand, vor etwa 4,55 Milliarden Jahren, bis zu der Zeit, wo man die ersten einzelligen Organismen findet, vor etwa 3,8 Milliarden Jahren, vergingen etwa 700.000 Millionen Jahre. In diesen 700.000 Millionen Jahre soll sich die erste lebende Zelle als der "gemeinsame Vorfahr alles Lebens auf Erde" von selbst aus lebloser Materie entwickelt haben. Aber für diesen Glauben gibt es überhaupt keinen sedimentären Beweis. In diesen 4 Milliarden Jahren, seit einzellige Organismen auf der Erde leben, haben sie sich überhaupt nicht in irgendwelche höhere Formen des Lebens entwickelt. Die erste lebende Zelle war schon ebenso komplex, wie diejenigen, die heute leben. Die chemischen Systeme, die sie damals benutzt haben, benutzen sie noch heute. Einige dieser ersten Zellen kann man nach ihrer Gattung und sogar nach ihrer Spezies bestimmen, von einzelligen Organismen, die heute noch leben. Die Zelle selbst hat überhaupt nichts "entdeckt" oder "erfunden". Sie hat sich überhaupt nicht in irgendwelche höhere Formen des Lebens entwickelt. Das ist nur eine fromme, religiöse Mythe. Das hat überhaupt nichts mit ernsthafter Wissenschaft zu tun.