Warum gibt es lebende Zellen? Warum können sie leben? Warum kann die Zelle ihre eigenen Bausteine herstellen? Wie kompliziert sind ihre chemischen Reaktionen? Wenn Sie, geehrter Leser, einige der Fachausdrücke nicht verstehen (oder sogar die meisten von ihnen), sollte sie das nicht beunruhigen. Ich möchte Ihnen hier nur zeigen, wie viel die winzige Zelle über Biochemie weiß und was sie kann. Dann fragen Sie sich bitte: Warum weiß dieser kleine Einzeller das alles über die molekulare Biologie und Biochemie, was ein Professor für molekulare Biologie und Biochemie zuerst lernen muss? Viele der technischen Details in der lebenden Zelle sind so kompliziert, dass die Experten nur einige verstehen. Einige verstehen sie nur zum Teil. Und einige, überhaupt nicht. Wenigstens bis jetzt noch nicht.
Stoffwechsel
Bruce Alberts ist ein amerikanischer Professor der Biologie. Er ist Präsident der Nationalen Akademie der Wissenschaften (NAS) von Amerika. Er und seine Mitarbeiter glauben, dass sich das Leben auf Erde von selbst entwickelt hat. Sie behaupten sogar in ihrem Lehrbuch The Cell (1989:41): "Es gibt heute keinen Platz für die Ansicht, dass es außerhalb der Gesetze der Chemie und Physik sonst noch irgend etwas gibt."
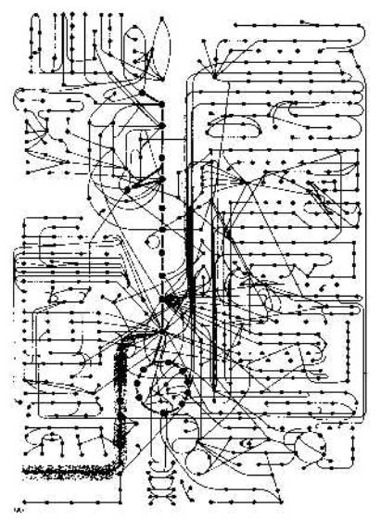
Aber Professor Bruce Alberts sagt dann: "Wie kompliziert eine Zelle ist, wenn man sie als eine chemische Maschine betrachtet, kann man erkennen, wenn man sich die Abbildung 2-35 (in seinem Buch) ansieht. Es ist eine graphische Darstellung, die nur einige der enzymatischen Pfade in einer Zelle zeigt. Alle diese Reaktionen entstehen in einer Zelle, die einen Durchmesser von weniger als 0,1 mm hat. Und für jede Reaktion braucht man ein Enzym. Dieses Enzym ist selbst in einer ganzen Folge von Informations-Übertragungen und Protein-Synthese-Reaktionen entstanden. Für ein typisches kleines Molekül - die Aminosäure Serin, zum Beispiel - gibt es ein halbes Dutzend oder mehr Enzyme, die es auf verschiedene Weise chemisch verändern können.
"Es kann mit AMP verbunden (adenylatiert) werden, in Vorbereitung für Proteinsynthese, oder abgebaut werden zu Glycin, oder umgewandelt werden zu Pyruvat, wenn es oxidiert werden soll. Es kann acetylatiert werden zu Acetyl CoA oder kann in eine Fettsäure umgewandelt werden, um Phosphatidyl-Serin zu machen. Alle diese verschiedenen Pfade möchten das gleiche Serin-Molekül haben. Und ähnliche Konkurrenz um Tausende andere kleine Moleküle gibt es zur gleichen Zeit. Man könnte meinen, dass das ganze System so fein eingestellt ist, dass irgendein kleiner Fehler, z.B., wenn es eine Zeit lang anderes Futter bekommt, katastrophal wäre.
"Doch die Zelle ist recht stabil. Jedes mal, wenn die Zelle gestört wird, versucht sie, ihren ursprünglichen Zustand wiederherzustellen. Sie kann sich anpassen und weiter funktionieren, während sie verhungert oder krank ist. Viele verschiedene Mutationen können bestimmte Reaktionspfade vernichten. Doch - solange bestimmte Mindestanforderungen erfüllt werden - überlebt die Zelle. Weil dann ein kunstvolles Netzwerk von Kontrollmechanismen die vielen Reaktionen koordiniert. ... Hier geht es uns jetzt nur um die einfachsten Mechanismen, die den Fluss der kleinen Moleküle durch die verschiedenen Stoffwechselpfade regeln." Alberts, B. et al. (1989:80).
Die Koordination von Katabolismus und Biosynthese in der Zelle. Nach Bruce Alberts et al. (1989:83) Abb. 2-35. Dies zeigt uns, wie kompliziert eine Zelle ist, wenn man sie als eine chemische Maschine betrachtet. Diese Tabelle zeigt nur einige der enzymatischen Pfade in einer Zelle. Alle diese Reaktionen kommen in einer Zelle vor, die weniger als 0,1 mm im Durchmesser ist. Und jede Reaktion erfordert ein Enzym. Und dieses Enzym ist aus einer ganzen Folge von Informations-Übertragungs- und Protein-Synthese-Reaktionen entstanden. Information entstammt immer einer nicht-materiellen, geistigen Welt. Sie kommt von einer intelligenten Person: vom Schöpfer.
Stoffwechselpfade regeln
Die Stoffwechselpfade in der Zelle werden geregelt. Wie?
Prof. Bruce Alberts: "Die Konzentration der verschiedenen kleinen Moleküle in einer Zelle ist gegen bedeutende Änderungen geschützt. Und zwar durch einen Vorgang, den man als Reaktions-Regulierung bezeichnet. Er stellt den Fluss der Metaboliten durch einen besonderen Pfad fein ein, indem er die Aktivität entscheidender Enzyme vorübergehend verringert oder beschleunigt. Das erste Enzym einer Folge von Reaktionen, zum Beispiel, wird normalerweise durch eine negative Reaktions-Wirkung des letzten Produktes dieses Pfades gehemmt: Wenn sich große Mengen des letzten Produktes ansammeln, wird ein weiterer Eintritt von Vorläufern im Reaktionspfad automatisch gehemmt. Wo sich die Pfade verzweigen oder überschneiden, wie das oft geschieht, gibt es normalerweise mehrfache Kontrollpunkte für verschiedene Endprodukte. Wie kompliziert solche Reaktions-Kontroll-Vorgänge sind, zeigt uns Abb. Zahl 2-37 (in seinem Buch). Sie zeigt uns das Muster der Enzym-Regulierung, die man in einem Satz verwandter Aminosäure-Pfade beobachtet hat.
"Die Reaktions-Regulierung kann fast unverzüglich funktionieren und ist auch umkehrbar; außerdem kann ein bestimmtes Endprodukt Enzyme aktivieren, die auf anderen Pfaden verlaufen. Und sie kann Enzyme hemmen, die ihre eigene Synthese verursachen." (1989:80).
Kontrolle von Auftank-Reaktionen (fueling reaction)
Wie kontrolliert die bakterielle Zelle ihre auftankenden Reaktionen? Warum weiß sie, welche Auftank-Reaktion richtig ist, und welche falsch ist?
Prof. F. C. Neidhardt und Mitarbeiter: "Allosterische Hemmung und Aktivierung spielen wichtige Rollen im Regulieren der Strömung durch Auftankpfade. Hier genügt das einfache Gerät der Endprodukt-Kontrolle beim ersten oder einem frühen Schritt in seiner Formation nicht mehr. Der Pfad der Formation der 12 Vorläufer-Metaboliten ist viel zu sehr miteinander verflochten (einige sind zyklisch), so dass jeder seine eigene Formation kontrollieren könnte, indem er einen bestimmten frühen Schritt hemmt. - Es gibt keine bestimmten frühen Schritte.
"Statt dessen arbeiten die Kontrollen innen in jedem Haupt-Auftankpfade (Abb. 4 in seinem Buch). Phosphoenolypyruvat, zum Beispiel, eine Zwischenform der Glycolyse, hemmt die Phosphofructokinase. Dieses Enzym, das eine Reaktion drei Schritte zurück im gleichen Pfad katalysiert. Und a-Ketoglutarate, eine Zwischenform vom Tricarboxyl-Säure Zyklus (TCA), hemmt die Citrat-Synthase, welche die Reaktion zwei Schritte zurück katalysiert.
"Man sollte bedenken, dass nicht alle Kontrollen der zentralen Auftank-Reaktionen durch allosterische Wechselwirkungen geschaffen werden können. Der entgegengesetzt Fluss der Strömung im Embden-Meyerhof-Parnas (EMP) Pfad, zum Beispiel, um die Gluconeogenese, während die Zellen auf Malat oder Pyruvat wachsen, zu bewirken, kann vorkommen. Weil alle, außer zwei der Reaktionen zwischen Pyruvat und Hexose-Phosphat, Gleichgewichts-Konstanten nicht weit weg von 1 haben. Und die beiden anderen, deren Fluss praktisch nicht umkehrbar ist, werden von zwei anderen Enzymen umgangen: von Phosphenolpyruvatesynthetase, das die Pyruvate-kinase umgeht, und eine bestimmte Phosphatase, die den Fruchtzucker-1,6-Diphosphat hydrolysiert.
"Um die Herstellungsmenge der 12 Vorläufer-Metaboliten unter einander auszugleichen und sie den Anforderungen der biosynthetischen Pfade anzupassen, braucht man diese Verbindungen als allosterische Effektoren, wie teilweise in Abb. 4 gezeigt. Aber das Bedürfnis für Koordination geht jenseits der Kohlenstoff enthaltenden Metaboliten. Die Auftank-Reaktionen müssen ATP liefern und müssen die Pyridine-Nukleotide reduzieren (NADH2 und NADPH2). Es überrascht deshalb nicht, dass diese Verbindungen allosterische Effektoren an mehreren Stellen in den zentralen Auftankpfaden sind." (1990:310, 311).
Wenn der Einzeller isst, wenn er sich repariert und wenn er sich verdoppelt, muss er gleichzeitig viele Dinge und in der richtigen Ordnung tun. Wie macht dieses winzige Geschöpf das?
Prof. F. C. Neidhardt und Mitarbeiter schreiben in ihrem Lehrbuch Physiology of the Bakterial Cell, unter der Überschrift "Regulierung des Genausdrucks: Multigene Systeme und Globale Regulierung":
"Bei vielen bakteriellen Aktivitäten müssen die Gene auf einer Organisationsebene koordiniert werden, die höher liegt, als die einzelnen Transkriptions-Einheiten. Die meisten, wenn nicht alle, Operone einer bakteriellen Zelle gehören zu einer Organisation, die auf höherer Ebene regelt wird. Wir nennen sie regulierende Netzwerke:
"Das Operon ist ein Kennzeichen der prokaryotischen Zelle. Diese Form der Organisation - bei der die Gene eines ganzen Pfades zu einer einzigen transkriptionellen Einheit verbunden werden - bewirkt eine einfache Lösung des Problems, wie man Gene mit verwandten Funktionen koreguliert. Warum? Muss die bakterielle Zelle über ihre offensichtlich erfolgreiche Strategie hinaus gehen?
Zu dieser Frage gibt es mindestens zwei Antworten. Die erste lautet: Bei einigen bakteriellen Vorgängen braucht man so viele Gene, dass man sie in einem einzigen funktionierenden Operon unterbringen muss. Das geschieht bei der Übersetzungsmaschinerie. Dort braucht man direkt eine Gruppe von mindestens 150 Genprodukten (ribosomale RNA, ribosomale Proteine, Einleitung, Verlängerung, und Schlussfaktoren, Amino-Acyl-tRNA Synthetase und tRNAs), um aus Aminosäuren Protein herzustellen.
"Sie sind so zahlreich, dass es schwierig, wenn nicht unmöglich wäre, ihre Synthese zu koordinieren, indem man ihre Gene in einen einzelnen Operon zusammen fügt. Doch die koordinierte Regulierung dieser Gene ist wichtig, damit sie richtig wachsen kann. ... Diese Koordination erreicht man, obwohl diese Gene in viele Dutzend getrennter Operonen organisiert sind.
"Die zweite Antwort lautet: Bei einigen bakteriellen Vorgängen braucht man einige Gene, die unabhängig von einander reguliert werden, und die von einer höheren Stelle aus koordiniert kontrolliert werden. Das sieht man am eindeutigsten bei der Gruppe von Genen, welche die katabolischen Enzyme codieren. Sie sorgen dafür, dass aus Zucker, Aminosäuren und anderen Verbindungen, Kohlenstoff und Energie entstehen. Wenn die Zelle in solch einer Umwelt lebt, die eine Mischung solcher Verbindungen enthält, verlangt es die Wirtschaftlichkeit, dass man nur eines der besten Substrate metabolisert, das am besten den Bedarf der Zelle an Kohlenstoff und Energie decken kann. (Für enterische Bakterien ist Traubenzucker so ein Substrat; für Pseudomonad ist es Succinat.) Die Operone, die die Enzyme für den Stoffwechsel der sekundären, überflüssigen Substrate verschlüsseln, sollten unterdrückt werden. Jeder Operon muss aber individuell veranlasst werden, wenn sein verwandtes Substrat in der Abwesenheit des besten Substrates anwesend ist. Diese doppelte Anforderung erfordert eine Organisationsebene, die über der des Operons liegt." (1990:351, 352).
Multigene regulierende Netzwerke
Wie viele multigene Systeme gibt es in einer bakteriellen Zelle? Wie arbeiten sie? Und wie komplex sind sie?
Prof. F. C. Neidhardt und Mitarbeiter: "Man schätzt, dass die bakterielle Zelle mehrere Hundert multigene Systeme entwickelt hat. Ihre Entdeckung und Analyse hat gerade erst angefangen. Einige der gut bekannten Beispiele für das enterische Bakterium Escherichia coli:
· Netzwerke, die reagieren, wenn von einem oder von mehreren Nährstoffen zu wenig vorhanden ist - Kohlenstoff und Energie, Ammoniak, anorganisches Phosphat,
· Netzwerke, die man bei Oxydations-Reduktions-Reaktionen und beim Elektronentransport braucht,
· Netzwerke, die reagieren, wenn durch Oxydation, Strahlung, hohe und niedrige Temperatur und extremen osmotischen Druck Schaden entstanden ist,
· Verschiedene Netzwerke, einige mit Unternetzwerken, die bedeutende Änderungen in der Physiologie und Morphologie der Zelle bewirken.
Die bakterielle Zelle hat verschiedene Wege entwickelt, individuelle Operon in koordinierte Netzwerke zu weben. Wie das geschieht, beginnt man erst jetzt etwas zu verstehen. In einigen Fällen hat man den allosterischen Proteinregler einfach von der Operon-Regulierung geborgt: ein Protein-Repressor oder Aktivator erkennt eine besondere Reihenfolge, die häufig in den Gebieten vorkommt, die von den Operonen kontrolliert werden. Diese Einrichtung gebraucht man in den SOS, Oxydations-Schaden, und anaeroben Elektron-Transport-Systemen in enterischen Bakterien.
"In anderen Systemen ist das Netzwerk ein alternativer Sigmafaktor. Es reprogrammiert die RNA Polymerase, um die Promotoren der anderen Operonen zu erkennen. Die Hitzeschock- und die Sporulationssysteme verschiedener bakterieller Arten veranschaulichen diese Situation. Bei anderen Netzwerken sind Proteinregler und Sigmafaktoren miteinander verbunden, wie in dem System vieler Bakterien, die Stickstoff verwenden. Eines der durchdringendsten Netzwerke, das strenge Operon, wird vom Nukleotid-Guanosin-Tetraphosphat (ppGpp) in einer Art reguliert, die man noch näher untersuchen muss.
"Viele regulierende Netzwerke hat man zuerst als zellulare Antworten auf Umweltänderungen erkannt. Deshalb kann die formelle Struktur eines Anreiz-Antwort-Systems die Masse von Informationen über jedes Netzwerk nützlich organisieren. Ein Anreiz-Antwort-System (Abb. 1 in seinem Buch) enthält einen Pfad. Er beginnt mit einem Anreiz aus der Umwelt. Es beeinflusst irgendein zellulares Ziel, oder einen Sensor, der ein Signal erzeugt. Dieses Anreiz-Antwort-System beeinflusst direkt oder indirekt, wenn das Signal durch einen oder mehrere Transducer (Signalumformer) geht, die Aktivität oder die Synthese von einem Regler, der die Ausgabe kontrolliert. Das ist normalerweise irgendeine adaptive Antwort auf Umweltänderung. Dieses System enthält oft einen Reaktions-Kontrollmechanismus, durch den die Zelle in den Prästimulus-Zustand oder in ein neues Gleichgewicht zurückkehren kann, das der veränderten Umwelt angepasst ist.
"In einem bakteriellen Gen-Netzwerk fließt ein Anreiz aus der Umgebung (zum Beispiel, veränderte Temperatur, Zustand des Futters oder Giftigkeit) in irgendeinen Sensor. Der erzeugt ein Signal, um die Aktivität oder die Synthese (normalerweise) eines Proteinreglers der anderen Gene zu beeinflussen. Die Produkte der anderen Gene sorgen dafür, dass die Zelle richtig wächst und überlebt. Modulatoren irgendeiner Art sorgen dafür, dass sie wieder in den Prästimulus-Zustand der anderen Operonen zurückkehrt oder schaffen ein neues Ausdrucksniveau.
"Es ist klar: viele der Operonen von E. coli gehören schon zu einem oder mehreren Netzwerken. Mit Hilfe des Rahmens, in Abb. 1, wollen wir jetzt einige dieser Netzwerke näher untersuchen, um die Prinzipien multigener Physiologie und molekularer Biologie zu veranschaulichen. Die ersten beiden Netzwerke, um die es hier geht, haben wir zum Teil gewählt, weil sie wegen ihrer Menge in der Zell-Physiologie wichtig sind. Zusammen kontrollieren sie wahrscheinlich, direkt oder indirekt, drei Viertel des Proteinaufbaus der bakteriellen Zelle." Neidhardt, F. C. (1990:352-357).
Kataboliten-Unterdrückung von Auftank-Genen: was bedeutet das? Und wie funktioniert das?
Prof. F. C. Neidhardt und Mitarbeiter: "Die meisten Bakterienarten enthalten viele Gene, die katabolische Enzyme verschlüsseln. Sie richten sich gegen ein Dutzend verschiedene Verbindungen von unterschiedlichem Nährwert. Das Wachsen auf einer einzelnen Quelle von Kohlenstoff und Energie - auf einem Substrat - erfordert ein relativ hohes zellulares Niveau der Enzyme. Sie müssen das Substrat metabolisieren und die katabolischen Produkte in die zentralen betankenden Pfade leiten. Alle Stoffwechselpfade in der Zelle fließen von den Metaboliten, die von diesen katabolischen Enzymen erzeugt worden sind.
"Die Bakterien scheinen es zu spüren, ob ein katabolischer Pfad in einem bestimmten Zustand richtig ist. Dann regeln sie den Genfluss dementsprechend. Wir haben schon erwähnt: Das Programm, das darüber entscheidet, welche Enzyme es machen soll und wie viele, muss höher entwickelt sein, als das Programm, das die Zelle einfach auffordert, die katabolischen Enzyme für Substrate herzustellen, die es in der Umgebung erkannt hat. Ein halbes Jahrhundert lang hat man angestrengt versucht, herauszufinden, wie das funktioniert. Doch das Programm der Zelle ist so kompliziert (so hoch entwickelt), dass die Physiologen die komplizierten Zusammenhänge einfach nicht verstehen können.
"Enterische Bakterien (die im Dickdarm von Säugetieren leben) verwenden mindestens vier verschiedene Vorgänge, um den vorrangigen Status von Traubenzucker als Kohlenstoff- und Energiequelle zu sichern:
· Konstitutive Synthese: Die Enzyme des Traubenzucker-Katabolismus sind in großem Maße ein Teil des EMP zentralen Auftankpfades. Davon werden viele hergestellt, ganz gleich, welche Substrate in der Umgebung vorhanden sind.
· Starter-Ausschluß: Traubenzucker verhindert den Eintritt bestimmter anderer induzierender Substrate durch Inaktivierung ihrer Permease.
· Vorübergehende Unterdrückung: Zusatz von Traubenzucker erzeugt eine vorübergehende, starke Hemmung der Synthese induzibler katabolischer Enzyme für etwa eine halbe Generation, auch wenn ihr Induzer (Starter) vorhanden ist.
· Katabolische Unterdrückung: Der Stoffwechsel von Traubenzucker erzeugt eine fortgesetzte, aber nicht so strenge Hemmung induzierter Enzymsynthese, sogar in der Gegenwart geeigneten Induzer.
"Von diesen Vorgängen versteht man bis jetzt noch keinen vollständig. Und die Kataboliten-Unterdrückung ist besonders komplex. Dazu gehört ein Element, über das man schon viel gelernt hat: das CAMP-CAP-Netzwerk. In E. coli gehört das lac-Operon zu diesem Netzwerk. Die gesamte Anzahl der Operonen kennt man nicht; aber es mögen mehrere Dutzend sein. Dazu gehören auch diejenigen, die mit dem Stoffwechsel von Galactose, Arabinose, Malzzucker, Tryptophan, D-Serin und Histidin zu tun haben. Die Operonen, die zu dieser Gruppe gehören, besitzen eine charakteristische Nukleotid-Reihenfolge, an die sich das regelnde Protein CAP (katabolisches Gen-Aktivator-Protein, ein Produkt des crp-Gens) bindet." Neidhardt, F. C. et al. (1990:357, 358).
Signal-Transduktion in Multigenen Systemen
Signal-Transduktion in multigenen Systemen, was ist das? Wie funktioniert das? Und wie komplex ist das?
Prof. F. C. Neidhardt und Mitarbeiter: "Das arc-Netzwerk mit den anderen drei Netzwerken, gerade beschrieben, behandeln ein gemeinsames Thema: Informationsverarbeitung durch Protein-Protein-Wechselwirkung. Schauen wir uns das etwas näher an.
"Die Verarbeitung physiologischer Information ist für uns kein neues Thema. Wir begegneten einigen Aspekten der Informationsverarbeitung, als wir die konformationellen Änderungen in bestimmten, (allosterischen) Proteinen betrachteten. Sie entstehen durch das umkehrbare Komplizieren bestimmter kleiner Moleküle. Die kleinen Moleküle - welche die Endprodukte biosynthetischer Pfade, von Startern katabolische Pfade, von Vorläufer-Metaboliten oder Nukleotiden sind - befördern Information (über ihre zellulare Konzentration) zum allosterischen Protein. Dieser Vorgang umfasst den größten Teil der Kontrolle der Enzym-Aktivität auf biosynthetischen Pfaden und beim Betanken von Reaktionen. Und auch das zentrale strategische Gerät der Operon-Physiologie - Aktivierung und Unterdrückung der Abschrift durch Proteine. In einer Anpassung können sie die DNA anbinden, und in der anderen können sie das nicht." (1990:375-377).
Das Regulieren von Multigenen Netzwerken
Wie reguliert die bakterielle Zelle ihre multigenen Netzwerke? Warum sind diese Wechselwirkungen wichtig?
Prof. F. C. Neidhardt und Mitarbeiter: "Protein-Protein-Wechselwirkungen sind beim Regulieren der Netzwerke, wie oben beschrieben, wichtig. Aber sie sind nicht die einzigen Mittel, mit denen man die multigenen Netzwerke steuern kann. Die molekularen Mittel durch die die verschiedenen Operon-Gruppen in ihrer Tätigkeit (Expression) koordiniert werden, ist ganz verschieden. Um diese Vielfalt zu verdeutlichen, betrachten wir drei Netzwerke, die einander in einem funktionellen Aspekt ähneln. Jedes wird veranlasst, stark zu reagieren, wenn sich etwas Bestimmtes in der Umwelt ändert. Doch der Mechanismus dieser Induktion ist in jedem Fall sehr verschieden.
"Wenn es heißer wird, schaltet sich in den meisten bakteriellen Zellen ein Satz von ungefähr 20 Genen ein. Das ist ihre Hitze-Schock-Antwort. Sie hat sich in einem unglaublichen Ausmaß erhalten: von den Archaebakterien und Eubakterien aufwärts durch die niedrigeren Eukaryoten zu den höheren Pflanzen und Tieren. Praktisch jede mikrobielle Zelle und jede Pflanze und tierische Zelle reagiert, wenn sich die Temperatur in ihrer Umgebung ändert. Wenn die Umwelttemperatur etwas höher liegt, als sie für diesen Organismus normal ist, stellt sie Hitzeschock-Proteine her.
"Noch ein Netzwerk - mit ungefähr der gleichen Anzahl von Genen - ist das SOS System. Diese Gruppe von Genen wird zu höherer Tätigkeit veranlasst, wenn die DNA der Zelle beschädigt ist, zum Beispiel durch UV Strahlung. Die Proteinprodukte dieser Gene arbeiten in verschiedenen Vorgängen, bei denen die beschädigte DNA repariert wird. Sie verhindern auch, dass sich die Zellen teilen, bis der Schaden repariert ist. Dieses Netzwerk schaltet sich ein, wenn sich die anderen Operonen proteolytisch spalten.
"Das lexA-Gen stellt den Repressor her, wenn die DNA einer Zelle beschädigt worden ist. Das RecA Protein wird aktiviert, um die proteolytische Spaltung von LexA zu erleichtern. Es schaltet dabei die LexA-gesteuerten Gene ein, die das SOS System bilden.
"Das oxyR Netzwerk arbeitet noch anders. Diese Gruppe eines Dutzends oder mehr Genen, zum Beispiel, schaltet sich bei oxidativem Schaden ein, der durch Wasserstoff-Peroxid verursacht wird. Diese Gene verschlüsseln schützende Enzyme, wie Catalase-Hydroperoxidase I und Alkylhydroxyperoxidase. Dieses Netzwerk schaltet sich ein, wenn der positive Reglers OxyR, ein Produkt des oxyR-Gens, aktiviert wird.
"Diese Beispiele zeigen uns: Die Regulierung der Gen-Netzwerke ist ebenso verschieden, wie die Regulierung individueller Operone." Neidhardt, F. C. (1990:380, 381).
Proteinsynthese: wie entstanden?
Warum macht die lebende Zelle Proteine? Und warum weiß sie, wie sie diese machen muss? Und warum übersetzt ein bestimmter Satz von Nukleinsäuren den Text in Proteine? Mit anderen Worten: Warum weiß ein bestimmtes Zeichen (oder Buchstabe) des DNA Codes, was es bedeutet? Und warum bedeutet es das?
Bruce Alberts und Mitarbeiter geben in ihrem Lehrbuch Molecular Biology of the Cell (1989:219) zu, obwohl sie an Evolution glauben: "Die molekularen Prozesse, auf denen die Proteinsynthese beruht, sind unerklärlich komplex. Wir können zwar viele von ihnen beschreiben. Aber konzeptionell ergeben sie keinen Sinn, wie zum Beispiel die DNA-Umschrift, DNA-Reparatur und DNA-Reproduktion. Die Proteinsynthese in heutigen Organismen beruht auf einer sehr großen Ribonukleoprotein-Maschine, dem Ribosom. Es besteht aus Proteinen, die den Kern der rRNA-Moleküle umgeben.
"Warum sollte es überhaupt rRNA-Moleküle geben? Und warum sind sie in der Struktur und der Funktion des Ribosoms so wichtig? Die Antwort hülfe uns, die Proteinsynthese zu verstehen. ... Die Proteinsynthese ist auch sehr auf eine große Anzahl von verschiedenen Proteinen angewiesen, die an die rRNAs im Ribosom gebunden sind. Dieser Vorgang mit so vielen verschiedenen Bestandteilen, die aufeinander angewiesen sind, hat dazu geführt, dass viele Biologen verzweifeln. Denn sie glauben nicht, dass sie jemals verstehen werden, wie die Proteinsynthese entstanden ist.
Wir mögen uns noch daran erinnern, was Professor Bruce Alberts und seine Mitarbeiter in ihrem Lehrbuch The Cell (1989:41) behaupteten: "Es gibt heute keinen Platz für die Ansicht, dass es außerhalb der Gesetze der Chemie und Physik sonst noch irgend etwas gibt."
Ist das logisch, ist das vernünftig, ist das wissenschaftlich? Überhaupt nicht. Die Behauptung:
"Es gibt heute keinen Platz für die Ansicht, dass es außerhalb der Gesetze der Chemie und Physik sonst noch irgend etwas gibt", ist nicht logisch, ist nicht vernünftig, ist nicht wissenschaftlich. Der Grund: Ein Gesetz kommt immer von einem Gesetzgeber. Die Gesetze der Chemie und Physik enthalten höchste wissenschaftliche Information und Mathematik. Menschliche Wissenschaftler hier auf der Erde haben bis jetzt nur einige von ihnen verstanden. Einige dieser Gesetze der Chemie und Physik haben sie nur etwas verstanden. Information und Mathematik existieren unabhängig vom Menschen. Der menschliche Wissenschaftler hat sie nur gefunden und sie etwas verstanden. Information und Mathematik sind etwas nicht-materielles, geistiges. Sie entstammen einer geistigen, nicht-materiellen Welt. Sie kommen vom wahren Gott der Bibel, dessen Name Jehova ist.
Schon auf der Ebene des Codes wissen wir, dass ihn eine intelligente Person erdacht und gemacht hat. Ein Code (oder Alphabet) ist etwas geistiges, nicht-materielles. Eine intelligente Person hat ihn zuerst erdacht. Sie muss zuerst bestimmte Zeichen auswählen (Buchstaben und so weiter) und entscheiden, was sie bedeuten sollen. Mit diesen Zeichen kann sie dann die Wörter, Sätze, Kapitel und das ganze Buch schreiben.
· Die 20 Aminosäuren sind die 20 Buchstaben des Aminosäure-Codes. Mit diesen 20 Buchstaben (und der Grammatik, die dazu gehört), erzeugt (schreibt) die lebende Zelle ihre Proteine.
· Die 4 Nukleinsäuren sind die 4 Buchstaben des Nukleinsäure-Codes. Mit diesen 4 Buchstaben (und der Grammatik, die dazu gehört), baut (schreibt) die lebende Zelle ihren genetischen Code, ihren DNA-Strang.
Die erste archaeelle Zelle auf der Erde war ebenso komplex, wie diejenigen, die heute leben. Sie hat überhaupt nichts "entdeckt" oder "etwas erfunden". Selbst der "einfachste" einzellige Organismus ist komplexer, als irgend etwas, was der Mensch bis jetzt erschaffen hat. Der schnellste heutige Supercomputer ist nur ein einfaches Kinderspielzeug im Vergleich zur einfachsten Zelle, die heute lebt, und die am Anfang auf dieser Erde, vor etwa 3,8-4,0 Milliarden Jahren, gelebt hat.
Die Hypothese der Evolution, oder des Neodarwinismus, wie jetzt allgemein in der Welt gelehrt, hat nichts mit ernsthafter Wissenschaft zu tun. Sie ist eine falsche Religion. Sie hat ihre Wurzeln in den Lehren des alten Babylonien (Sumer-Akkad) und Ägypten. Ihre heidnischen Priester lehrten schon vor etwa 3.500 bis 4.000 Jahren, das Leben auf der Erde habe sich von selbst aus dem Wasser und Schlamm des Euphrat und Tigris und des Nil entwickelt.