Grampositive und gramnegative Bakterien. Ihre Membran und Wand (Hülle). Wie sie ihre Wand machen. Wie sie sich voneinander unterscheiden. Was braucht man, um sie zu erdenken und sie zu machen? Beweist ihre Zellwand, dass sie alle von einem gemeinsamen Vorfahren abstammen?
Prof. Frederic C. Neidhardt und Mitarbeiter schreiben in ihrem Lehrbuch Physiology of the Bacerial Cell (1990:25-27) über die Zellwand der Bakterien: "Die Bakterien kann man in zwei breite Klassen einteilen: in grampositive und gramnegative. Grampositive und gramnegative Bakterien unterscheiden sich in einer Anzahl wichtiger Aspekte, außer in der Struktur der Hülle. Der grampositive-gramnegative Unterschied ist tiefgründig. Er spiegelt nicht nur einen fundamentalen Unterschied in der Wandstruktur wider, sondern auch andere wichtige biochemische, physiologische, und genetische Unterschiede. Die meisten, aber nicht alle, Bakterien, gehören diesen beiden Gruppen an. Diejenigen, die nicht zu diesen Gruppen gehören, gehören zu zwei anderen Klassen. Eine Gruppe ähnelt sehr den anderen Bakterien. Doch sie haben überhaupt keine Zellwand: die Mollikuten. Die andere Gruppe, die Archaebakterien, haben Zellwände. Doch die enthalten kein typisches Murein. Und sie unterscheiden sich auch in vielen anderen Dingen von der typischen prokaryotischen Zelle." (1990:25-27).
Zellwand
Was haben die Wissenschaftler jetzt über die Zellwand herausgefunden? Wie komplex ist sie? Was muss man alles wissen, damit man sie versteht und sie machen kann?
Prof. F. C. Neidhardt und Mitarbeiter: "Die Bakterienzellen sind besonders zäh. Das heißt, es ist schwierig, sie durch mechanische Mittel zu zerbrechen. Und sie behalten ihre Form sogar unter harschen Zuständen. Die Bakterien sind vor allem deshalb so zäh, weil sie eine Zellwand haben. Sie gibt ihnen einen starren mechanischen Halt. Und sie verhindert, dass der Wasserdruck die Zellen zerbricht (osmotische Lyse). Die Wand (Hülle) schützt die Zelle außerdem chemisch und physisch vor schädlichen Chemikalien, die der Zellmembran schaden könnten. Grampositive und gramnegative Bakterien unterscheiden sich beträchtlich in der Struktur ihrer Zellwände, und in der Art und Weise, wie sie gebaut sind. Sie helfen ihnen, mit ihrer sich ändernden Umwelt zurechtzukommen. Die Wand (Hülle) der grampositiven Zelle sieht wie eine dicke Decke aus. Die der gramnegativen Zelle, wie ein dünnes Laken. Doch in Wirklichkeit ist auch die dünne gramnegative Wand sehr zugfest." (1990:33).
Grampositive Wand
Prof. F. C. Neidhardt und Mitarbeiter: "Die grampositive Zellwand besteht aus einem dicken multimolekularen Mantel aus Muerein (eine Form von peptidoglykischen Polymeren). In ihm sind kleinere Mengen anderer Polymere verteilt, vor allem teichonische Säuren. Der polymere Stoff besteht aus vielen Schichten, die um die Länge und die Breite der Zelle gewickelt sind. Dadurch bilden sie einen Beutel, der die Größe und die Form des Organismus bestimmt. Das Murein kann man als eine Struktur intakt absondern. Das ist dann der Murein-Sack. Die Form des Bakteriums hängt von der Form des Sackes ab. Die meisten Bakterien sehen wie Stäbe aus (Bazillus) oder Helixen (Spirochät); einige sehen aus wie Spindeln, Seesterne oder flach gemachte Vielecke; andere haben noch andere, komplexere Formen. ... Das starre Murein-Netzwerk erlaubt es den Bakterien, in einer Umwelt zu überleben, die gewöhnlich hypotonisch ist. Das heißt, sie haben einen niedrigeren osmotischen Druck, als der ihres Cytoplasmas. Ohne diese starre, korsettähnliche Hülle, gegen die man drücken kann, würde die Membran zerplatzen und die Zelle auseinander fließen (lysen)." (1990:33-36).
Gramnegative Zellwand und die äußere Membran.
Prof. F. C. Neidhardt und Mitarbeiter: "Gramnegative Bakterien haben das Problem, wie man die cytoplasmische Membran schützt, ganz anders gelöst. Ihre Mureinschicht ist viel dünner, als die der grampositiven Bakterien. Und sie haben etwas ganz anderes gemacht - eine äußere Membran. Sie bauen sie außerhalb einer dünnen Mureinschicht auf. Die äußere Membran ist von den üblichen biologischen Membranen chemisch verschieden. Sie kann schädlichen Chemikalien widerstehen Das ist eine zweischichtige Struktur. Und ihr inneres Blättchen (leaflet) ähnelt in seiner Zusammensetzung der der cytoplasmischen Membran.
"Ihr äußeres Blättchen (leaflet) hat andererseits einen einmaligen Bestandteil an Stelle der Phospholipiden. Dieser Bestandteil ist das bakterielle Lipopolysaccharid oder LPS. Das ist ein komplexes Molekül. Und man findet es in der Natur sonst nirgendwo. Deshalb sind die Blättchen dieser Membran äußerst asymmetrisch. Und die Eigenschaften dieser außergewöhnlichen Doppelschicht unterscheiden sich beachtlich von denjenigen einer regulären biologischen Membran. Die Fähigkeit, hydrophobe (wasserabweisende) Verbindungen auszuschließen, ist unter biologischen Membranen außergewöhnlich.
"Das gramnegative Bakterium und grampositive Bakterium schließen hydrophobe Verbindungen aus, indem sie sich mit hydrophoben Polysacchariden umgeben. Aber diese unterscheiden sich in Struktur und Organisation in den beiden Gruppen. Die äußere Membran führt offenbar in eine Sackgasse. Wegen ihrer lipoiden Natur kann man erwarten, dass sie auch hydrophile (wasserfreundliche) Verbindungen ausschließt. In diesem Fall könnten keine Verbindungen, hydrophobe oder hydrophile, die äußere Membran durchqueren.
"Die gramnegativen Bakterien scheinen zwar das Problem gelöst zu haben, wie man die cytoplasmatische Membran schützt. Doch dabei scheinen sie ein neues geschaffen zu haben. Wie transportieren diese Organismen ihre Nährstoffe? (Wie nehmen sie diese auf?) Kopieren sie in der äußeren Membran die aktiven Transportgeräte der cytoplasmischen Membran? Diese Strategie wäre nicht nur verschwenderisch, sondern dadurch könnte auch die äußere Membran ebenso empfindlich gegenüber Umwelteinflüssen werden, wie die cytoplasmische Membran. Auch hier haben die Bakterien eine interessante Lösung gefunden: Die äußere Membran hat besondere Kanäle. Durch diese Kanäle kommen die hydrophilen Verbindungen wie Zucker, Aminosäuren, und bestimmte Ionen in die Zelle hinein. Diese Kanäle bestehen aus Proteinmolekülen. Man bezeichnet sie als Porine (= von Poren). Porin-Kanäle sind eng, gerade so groß, damit Verbindungen, die kleiner als 600-700 Dalton sind, dort hindurch können (Abb. 9 in ihrem Buch).
"Die Kanäle sind so klein, dass hydrophobe Verbindungen nicht durch sie hindurch kommen und draußen bleiben müssen. In gewissem Maße kann E. coli Porine verschiedener Kanalgrößen einbauen, um sich so den osmotischen Verhältnissen der Umwelt anzupassen.
"Einige hydrophile Verbindungen, welche die Zelle braucht, um zu überleben, sind größer als der Kanal in diesen Porinen. Zu diesen großen Molekülen gehören Vitamin B12, Zucker, die größer als Trisaccharide sind, und Eisenchelate. Solche Verbindungen durchqueren die äußere Membran durch getrennte, dafür vorgesehene Durchgangsmechanismen. Sie verwenden Proteine, die man speziell dafür vorgesehen hat, dass sie alle diese Verbindungen dort hindurch bringen. So erlaubt die äußere Membran den Durchgang kleiner hydrophiler Verbindungen, schließt hydrophobe Verbindungen, große oder kleine, aus und erlaubt den Eintritt einiger größerer hydrophiler Moleküle durch besonders dafür vorgesehene Mechanismen." (1990:37-40).
"Die äußere Membran ist keine völlig getrennte Struktur. Sie ist mit der Mureinschicht und der cytoplasmischen Membran verbunden. Die Verbindung mit der Mureinschicht wird von zwei Arten von Wechselwirkungen bewirkt. Die wichtigste ist ein äußeres Membran-Lipoprotein. Dieses Protein ist in etwa 700.000 Kopien je Zelle vorhanden. Es ist das zahlreichste Protein (nach seiner Anzahl, nicht nach seinem Gewicht) in E. coli.
"Etwa ein Drittel der Lipoprotein-Moleküle ist kovalent mit dem Murein zusammengefügt. Sie helfen mit, die beiden Strukturen zusammenzuhalten. Die andere Art der Wechselwirkung ist der feste (aber wahrscheinlich nicht kovalente) Verband einiger der äußeren Membran-Porine mit dem Murein. Zusammen stellen Lipoproteine und Porine in jeder Zelle über 400.000 verstrebte Kontakte zwischen der äußeren Membran und der Zellwand her." (1990:41).
Isst bestimmte Substrate
Welche Substrate kann das Bakterium E. coli aufnehmen? Wie nimmt es sie auf, da es keinen Mund hat?
Prof. F. C. Neidhardt und Mitarbeiter: "Fast alle Transportmechanismen von E. coli (auch das erleichterte Durchdringen, schockempfindlicher aktiver Transport, und sekundärer aktiver Transport) beteiligen sich am Aufnehmen von einem der Zucker oder Zuckeralkohole (Abb. 4 in ihrem Buch). Interessanterweise erfolgt die Aufnahme eines bestimmten Zuckers keinem Muster: E. coli, zum Beispiel, nimmt die Lactose durch Protonen-Transport auf, Staphylococcus aureus aber durch ein PTS.
"Aminosäuren werden von schockempfindlichen oder sekundären aktiven Transportsystemen transportiert. In E. coli bringen 14 verschiedene Transportsysteme die Aminosäuren in die Zelle. Man hat die Aminosäure-Transportsysteme der Bakterien untersucht. Dabei hat man mehrere Muster erkannt. Einige Systeme transportieren eine Gruppe von Aminosäuren mit ähnlicher Struktur. Oft haben diese Teilsysteme für nur eine der Aminosäuren. Diese offenbare Redundanz dient einem Zweck. Eines der Systeme hat eine hohe Affinität (Affinitäts-Konstante 10mal oder noch größer) und hohe Strömung. Jedes hat offensichtliche Vorteile in bestimmten Umgebungen. Außer den Aminosäure-Transportsystemen hat E. coli verschiedene Transportsysteme für Kofaktoren, verschiedene Ionen und Stoffwechsel-Zwischenstufen ". (1990:182, 183).
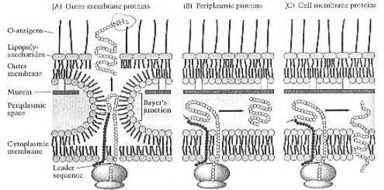
Wie ein Gram-negatives Bakterium sein eigenes Zellwand-Protein macht. Aus: Neidhardt et al. (1990:114) Bild 8. Die einfache Zelle kann etwas, was der Mensch auch am Anfang dieses 21. Jahrhunderts noch nicht geschafft hat: automatische Montage, automatische Produktion. Deshalb muss derjenige, der diesen einzelligen Organismus erdacht und erschaffen hat, viel mehr über Wissenschaft und Technik wissen, als irgendein Wissenschaftler, der jetzt auf der Erde lebt: der Schöpfer, dessen Name Jehova ist.
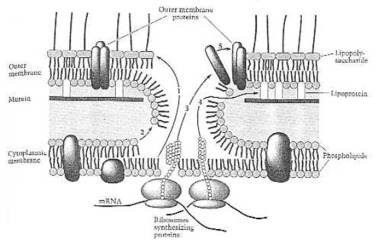
Montage-Ablauf in der äußeren Membran (Hautschicht). (1) Lipopolysaccharide;(2) Phospholipide; (3) äußere Membran-Proteine; (4) Lipoprotein (Fett); (5) Selbstmontage der äußeren Membran. Nach: Neidhardt et al. (1990:121) Bild 14.
Osmo-Regulierung und Porin-Ausdruck
Wie wird E. coli mit den Extremen des osmotischen Druckes in seiner Umwelt fertig? Und wie macht es seine Porine, die Poren in seiner äußeren Membran?
Prof. F. C. Neidhardt und Mitarbeiter: Die Bakterien müssen unter anderem mit dem osmotischen Druck in ihrer Umwelt fertig werden. Die bakterielle Zelle verwendet mehrere molekulare Strategien, um den Wirkungen gesundheitsschädlicher Extreme des osmotischen Druckes in ihrer Umwelt entgegenzuwirken.
"Kanäle durchqueren die äußere Membran. Sie sind von Proteinen umgeben, die wir als Porine bezeichnen. In E. coli gibt es zwei bedeutende Porine - OmpF und OmpC. Sie ergänzen sich gegenseitig. Die Poren, die OmpC macht, sind etwas kleiner. E. coli macht sie, wenn der osmotische Druck und die Temperatur sehr hoch sind. Dieses Muster bedeutet: das OmpC ist die vorherrschende Form, während die enterischen Bakterien im Darm wachsen. Es ist physiologisch sinnvoll, dass enterische Bakterien kleinere Poren haben, wenn sie im Darm leben, und dass sie größere haben, wenn sie draußen leben. Die kleineren Poren sorgen dafür, dass viele der giftigen Moleküle, die im Darm leben, nicht in die Zelle hinein kommen. Die größeren Poren erlauben es Substraten in verdünnten Lösungen, leicht in Zellporen hinein zu kommen. Weil gelöste Stoffe durch die äußeren Membran-Poren herein kommen müssen. Und wie schnell sie durch die Poren hindurch kommen, hängt davon ab, wie groß deren Durchmesser ist, und vom Konzentrations-Unterschied in der Membran.
"Die Gene, welche die bedeutenden Porine verschlüsseln (codieren), und bestimmte andere Gene, die äußere Membran-Eigenschaften beeinflussen, werden von zwei Proteinen gesteuert - von Produkten des zusammengefügten envZ (für envelope proteins) und ompR (für outer membrane protein regulation) Genen. Das EnvZ-Protein ist ein cytoplasmisches Membran-Protein. Man nimmt an, dass es als ein Osmo-Sensor funktioniert. Es ähnelt sehr bestimmten Protein-Kinasen. Weil sie sich so ähneln und aus anderen Gründen, und vom Phänotyp der Unsinn- und anderen Mutationen in envZ, glauben Forscher, dass dieser EnvZ mit dem zweiten Protein, OmpR zusammen arbeitet. Es beeinflusst seine Aktivität, indem es dieses phosphorylisiert.
"Das Protein OmpR verbindet sich besonders mit der DNA vor den Promotoren, die es steuert. Die Verbindungsstelle ist etwa -50 im ompF-Promotor, und -90 im ompC-Promotor. Ein aktuelles Modell dafür, wie man den alternativen Ausdruck von ompF erreicht, sieht so aus (Abb. 7 in ihrem Buch): Hoher osmotischer Druck phosphoriliert EnvZ. Und dieses aktivierte Protein phosphorylesiert dann OmpR. Die Phospho-OmpR (OmpR-P) unterdrückt die Umschrift von ompF, indem es sich an einer Stelle zwischen -40 und -60 im ompF-Promotorgebiet anbindet. Es aktiviert die ompC-Abschrift, indem es sich an Stellen mit niedriger Affinität zwischen -75 und -105 im ompC-Promotor-Gebiet anbindet. Wenn der osmotische Druck niedrig ist, ist das Niveau von OmpR-P niedrig. Und deshalb kann es dann ompC nicht aktivieren. Aber es kann immer noch ompF aktivieren, indem es sich an Hochaffinitäts-Stellen im -60 bis -100 Gebiet des ompF-Promotors anbindet.
"Die Wirkungen von OmpR-P, als ein Stimulator und als Schutzmittel von Abschriften dieser beiden Gene, wird noch durch einen anderen Mechanismus ergänzt. Das Gen micF wird in der entgegengesetzten Richtung von ompC von einem Promotor abgeschrieben, in der Nähe von ompC; micF verschlüsselt eine 174-Basen RNA, es ist komplementär zu einem Gebiet in der Nähe vom Anfang der ompF-Mitteilung. Die micF-Abschrift blockiert die Tätigkeit von ompC. Die Steuerung der micF-Tätigkeit ähnelt der des ompC. Deshalb bewirken die Zustände, die ompC einschalten, dass sie gleichzeitig micF einschalten. Dadurch helfen sie mit, ompF zu unterdrücken (Abb. 7 in ihrem Buch).
"Es gibt ein Netzwerk von Genen außer denjenigen für die beiden Porine, die vom envZ/OmpR-Paar interaktiver Proteine gesteuert werden; einige dieser Gene werden eingeschaltet, andere unterdrückt. Worum es physiologisch gesehen bei den Genen in diesem System geht, weiß man noch nicht. Die EnV/OmpR Geschichte zeigt uns, dass sie ein wichtiges Muster in der Protein-Protein-Wechselwirkung in Anreiz-Antwort-Netzwerken ist, auch in vielen, bei denen man noch nicht weiß, wie sie physiologisch funktionieren." - Neidhardt, R. C. et al. (1990:272, 373).
Wie nimmt die Zelle Futter auf? Und wie scheidet sie Abfall aus ihrem Körper aus, da sie doch keinen Mund und keinen After hat? Durch Schleusen in ihrer Zellwand. Der Aufbau und die Funktion dieser Zellwand sind so kompliziert, dass kein Mensch das völlig versteht, geschweige denn nachbauen kann. Aus: F.C. Neidhardt et al. (1990:178) Bild 1. Die Hülle (Haut) der bakteriellen Zelle entsteht immer nur aus einer schon bestehenden Hülle. Sie kann nicht neu gemacht werden. Die Hülle (Haut) der bakteriellen und archaeellen Zelle ist so kompliziert, dass kein menschlicher Wissenschaftler sie machen kann. Die Behauptung, sie habe sich von selbst entwickelt, ist reines Wunschdenken. Für diesen Glauben gibt es keine experimentellen und paläontologischen Beweise.
![]()
Wie die Zelle E. coli Zucker und Zucker-Alkohole aus ihrer Umwelt durch die Zellwand aufnimmt. Nach: Neidhardt et al. (1990:182) Bild 4.
Erste Zellmembran
Wie ist die erste Membran entstanden? Konnte sie von selbst in der chemischen Ursuppe entstehen?
Prof. Siegfried Scherer: "Ein Hauptbestandteil von Biomembranen sind Phospholipide, bestehend aus Glycerin, Phosphorsäure und langkettigen aliphatischen Verbindungen (welche mit Glyzerin als Ether oder Ester verknüpft sind). Diese Moleküle weisen wie alle grenzflächenaktiven Substanzen (Tenside) einen hydrophoben (= wassermeidenden) und einen hydrophilen (= wasserverträglichen) Bereich auf. Moleküle mit diesen Strukturmerkmalen können sich spontan zu Aggregaten (z.B. Doppelschichten, Mizellen, Vesikel) zusammenlagern. Synthesemöglichkeiten solcher Substanzen unter präbiotischen Bedingungen sind unbekannt.
"Biologisch aktive Membranen, wie wir sie von lebenden Zellen kennen, entstehen immer aus bereits existierenden und werden nicht de novo synthetisiert. Zu den Aufgaben von Biomembranen gehört nicht nur die Abgrenzung der Zelle gegen die Umgebung, sondern unter anderem auch der Stofftransport durch die Membran hindurch. Der komplexe Aufbau von Biomembranen spiegelt die vielfältigen Aufgaben (wie z.B. Erkennungsmechanismen, Zell-Zell-Kommunikation) von Zellmembranen wieder." (1998:146).
Hans Kleinig ist Professor für Zellbiologie und Ordinarius an der Universität Freiburg. Uwe Maier ist Professor für Zellbiologie an der Philipps-Universität Marburg. Sie schreiben in ihrem Lehrbuch Zellbiologie (1999:73) über die Biosynthese von Membranen: "Biomembranen können nicht de novo im Cytoplasma einer Zelle entstehen. Ort der Synthese von Membranbausteinen, Proteinen wie Lipiden, sind im allg. Membranen selbst: in Protocyten die Plasmamembran..."
Ergebnis
Biologisch aktive Membranen, wie wir sie von lebenden Zellen kennen, entstehen immer aus schon bestehenden Membranen. Sie werden nicht neu zusammengefügt. Wie solche Substanzen unter präbiotischen Zuständen entstanden sein könnten, weiß man nicht.
Wir haben uns kurz grampositive Zellen und gramnegative Zellen angeschaut. Die grampositive Zelle hat eine Wand. Die gramnegative Zelle hat eine Membran. Grampositive Zellen, gramnegative Zellen und Archaeabakterien unterscheiden sich sehr von einander. Sie sind überhaupt nicht mit einander verwandt. Sie sind ebenso von einander verschieden, wie die Pflanzen von den Tieren. Und nichts beweist, dass sie von einem gemeinsamen Vorfahren abstammen. Schon vor etwa 3,5 bis 3,8 Milliarden Jahren, am Anfang der Erdgeschichte, hat es grampositive und gramnegative Zellen und Archaebakterien gegeben, genauso wie heute.
Viele der Gattungen und Arten, die man bei Warrawoona, NW Australien gefunden hat, leben heute noch. Diese Einzeller waren ebenso komplex wie diejenigen, die heute leben. Sie haben sich überhaupt nicht in höhere Lebensformen entwickelt. Sie sind vollautomatische chemische Fabriken, mit einem Durchmesser von einem winzigen Bruchteil eines Millimeters. Wie entstehen chemische Fabriken und Häuser mit ihren Mauern und Dächern heute? Ein Haus (oder eine chemische Fabrik) entsteht zuerst im Verstand eines intelligenten Menschen.
· Zuerst hat er ein Bedürfnis. Er könnte sich sagen: "Ich möchte es warm und trocken haben, wo die Bären und Wölfe nicht hinkommen und mich töten können.
· Dann hat er einen Wunsch: "Ich möchte ein Haus (oder ein Zelt, oder eine Höhle) haben."
· Dann macht er einen Plan: "Mein Haus soll so und so groß sein. Es soll so und so viele Zimmer, Türen und Fenster haben. Und ich werde es mit Ziegeln (oder Steinen oder Baumstämmen) bauen."
· Dann benutzt er seinen Bauplan (Entwurf), Energie und das Baumaterial und baut das Haus.
Die ersten drei Schritte, die zum Haus führen, sind rein geistig, nicht-materiell. Ein intelligenter Mensch, als Baumeister ausgebildet, hat es zuerst erdacht. Sogar die Ziegelsteine im Haus (ihre Form und Funktion), hat ein intelligenter Mensch zuerst erdacht. Die Atome, aus denen die Ziegelsteine, (Steine und Baumstämme) bestehen, wissen nichts von einem Haus.
Materialisten haben behauptet: Alles kommt aus der Materie. Aber ein Haus wächst nicht aus den Steinen oder den Baumstämmen der sibirischen Taiga hervor, wie ein Baum. Warum nicht? Weil das Baumaterial des Hauses nichts über die Form und die Funktion eines Hauses weiß. Das alles kommt zuerst von einem intelligenten Menschen, der das erdacht hat.
Die Atome in der Zellwand (oder Membran) des Bakteriums sind auf seinem niedrigsten organisatorischen Niveau. Sie sind nur ein Mittel zum Zweck, wie die Ziegelsteine im Haus. Ein Bakterium ist sehr viel komplizierter als irgendein Haus, das der Mensch bis jetzt gebaut hat. Der Mensch kann ein Haus (und eine chemische Fabrik) bauen. Aber er kann keine lebende Zelle bauen, eine voll automatisierte chemische Fabrik. Das ist zu kompliziert. Das ist viel komplizierter, als irgend etwas, was der Mensch bis jetzt erschaffen hat. Die ersten lebenden Zellen auf Erde, vor etwa 3,5-4 Milliarden Jahren, waren ebenso kompliziert wie diejenigen, die heute leben.
Die Zelle weiß viel über Biochemie. Sie enthält Information und Mathematik. Aber die Zelle selbst hat nichts entdeckt oder erfunden. Information und Mathematik sind etwas Nichtmaterielles, Geistiges. Sie entstammen einer geistigen, nicht-materiellen Welt. Deshalb muss auch das Bakterium zuerst von jemandem erdacht und gemacht worden sein. Es hat sie jemand erschaffen, der viel mehr über Wissenschaft weiß, als irgendein Mensch, der jetzt auf der Erde lebt: der Schöpfer, der Gott der Bibel, dessen Name Jehova ist.