
We shall study in this volume the first living cells on earth. And we shall ask ourselves: How have they come into being? Why have they arisen? How complex has the first living cell on earth been? How large was it? What did it eat? How did it multiply? In what kind of a climate has it lived? What did it have to "know" about chemistry, physics, mathematics, and information processing, in order to live, to survive, and to become many? Do the oldest bacteria in the fossil record prove the philosophy or religion of evolution, of neo-Darwinism?
First, we shall look briefly at the sediments of the Early Precambrian and Late Precambrian Time. We shall look then at one-celled organisms, thriving at the limits of life: in hot or even boiling water. We shall also try to find out: How do these tiny creatures become many? How do some of them use the light of the sun as a source of energy? How they build their cell wall. Some of these one-celled organisms even have a rotary motor. It is the world's smallest rotary motor. How does the cell make it? And how does it work?
Since when has there been life on earth? When has the first living cell on earth arisen? Where? What are now the remains of the world's oldest cells? How large and how complex was the first cell? What was it able to do chemically? What did it eat? What would one have to know and to be able to do now, if one wanted to make such a first cell? Does a living cell contain any information, planning, and design? If so, why? How much must it know about biochemistry and biophysics, in order to live? Why does the bacterium know something, which a human scientist first has to learn?
Do the sediments of the Early Precambrian Time prove that all life on earth has evolved by itself from the "first cell", that it has come from a common ancestor? How different is today's cell from that of the beginning? Has the first cell evolved since then into higher forms of life, into plants, animals, apes, and human beings? Were the first living cells on earth primitive? And are there any primitive cells now? What have some of the world's foremost scientists found out about this now?
J. William Schopf
J. William Schopf, Dept. of Earth and Space Sciences, Molecular Biology Institute, University of California at Los Angeles, California, U.S.A., believes in evolution. He writes in Major events in the history of life (1992:37) under "Precambrian Microfossils":
"Precambrian microfossils occur principally in two types of sedimentary deposits: in cherts, rocks composed of minute interlocking grains of silica, occurring as the mineral quartz (SiO2), that have been deposited chemically, petrifying microscopic organisms in the place in which they live (for example, in the layers of stromalites); and in shales, rocks formed by consolidation of layers of clay or mud disseminated, along with phytoplankton and other debris, at the bottom of lakes or ocean basins. Although commonly twisted or otherwise distorted, the microfossils of chert deposits are generally unflattened, composed of three-dimensionally preserved organic-walled cells that are thoroughly embedded in, and infilled by, the petrifying fine-grained quartz. In contrast, the carbonaceous microfossils of shales have been preserved by compression, flattened between thin layers of consolidated silt."
Sometimes, the shale and chert have not preserved the shape of the living cell. Can one still find out then, if one-celled organisms have once lived there? If so, how?
Prof. J. William Schopf: "All living systems are composed of organic compounds, chemical combinations of the elements carbon (C), hydrogen (H), oxygen (O), nitrogen (N), and sometimes sulfur (S) and phosphorus (P). - 'CHONS.' This carbonaceous organic matter is commonly preserved in ancient rocks, coals, for example, are the compressed remnants of ancient plant debris. Thus it seems reasonable to suppose that the mere presence of carbonaceous matter in Precambrian sediments constitutes firm evidence of the existence of life. ... The cardinal question is, 'What is the biological source of this ancient carbonaceous matter - what type(s) of living systems originally made it?'
"Fortunately, the biochemical organization of living systems provides the means to answer this question. Organisms are not simply haphazard aggregates of CHONSP-type organic compounds. On the contrary, they are finely tuned organic systems, powered by energy supplied from external sources (for example, sunlight) and organized internally by an ordered, complex network of biochemical reactions. These reactions, in turn, are facilitated, that is, they are catalyzed, by protein-type compounds known as enzymes, with each biochemical reaction catalyzed by a specific enzyme.
"The first biochemical step in both anoxic and oxygenic photosynthesis, for example, is a reaction resulting in fixation of the carbon atom of carbon dioxide into an organic compound. Both in photosynthetic bacteria and in cyanobacteria, this reaction is catalyzed by an enzyme known as fibulose bisphosphate carboxylase/oxygenase or, in shorthand Rubisco. Because of the unique properties of the Rubisco enzyme, it leaves a tell-tale isotopic signature in its products, a signature that can be deciphered in organic matter having an age even as great as 3 500 Ma." Schopf, J. W. (1992:38, 39).
How does that work?
Prof. J. W. Schopf: "In nature, carbon atoms exist in three different forms, or isotopes, differentiated from each other by their subatomic structure. One of these isotopes, carbon-14 (written in shorthand as 14C), is radioactive. Because it is unstable, disintegrating over time, it cannot be detected in materials older than about 50,000 to 60,000 years.
"The other two isotopes of carbon, 13C and 12C, are both stable; neither disintegrates over time. Thus, throughout all of earth history, two types of carbon dioxide have existed in the atmosphere, 13CO2 and 12CO2. The carbon-fixing enzyme in photosynthesis, Rubisco, has the notable property of discriminating between these two types of carbon dioxide, preferentially tending to react with 12CO2 and, therefore, to fix preferentially the isotopically lighter stable carbon isotope, 12C, into the organic compounds produced.
"Thus, both in photosynthetic and in cyanobacteria (and in all other photoautotrophs as well), the products of Rubisco-catalyzed photosynthesis are enriched somewhat in 12C relative to the concentration of this isotope in the atmospheric CO2. Like health-food stuffs, 'photoautotrophs eat light' (both sunlight and isotopically light carbon)!
"Atmospheric carbon dioxide is also involved in inorganic chemical processes: CO2 dissolved in oceanic waters is converted to bicarbonate (HCO3-), which can react with calcium to form calcium carbonate, CaCO3-), the mineral material of which limestones are composed. As a result of these inorganic processes, and in contrast with the 12C-enriched products of photosynthesis, the carbonate carbon in limestones is depleted slightly in 12C relative to the atmospheric CO2 source.
"Although the amounts of isotopic discrimination resulting from these various processes are small, they can be measured easily by use of a mass spectrometer. In typical present-day environments, biologically produced organic carbon is enriched in 12C by about 17 parts per thousand (17 %o), relative to atmospheric CO2, whereas the inorganic, carbonate carbon of limestone is depleted in 12C by about 7%o, a net difference between the two types of carbon of 24%o. The amount of isotopic discrimination can vary depending on environmental conditions, such as the amount of CO2 in the earth's atmosphere, a concentration that is thought to have changed, probably quite considerably, over the history of the planet. Nevertheless, a net difference of about 20 %o to more than 40 %o between the isotopic composition of biological and inorganic carbon can be traced far into the geologic past. Indeed, this isotopic signature has been detected in hundreds of Precambrian rock samples, the oldest some 3 500 Ma in age. Together with studies of Precambrian stromatolites and microfossils, these biogeochemical data provide important evidence regarding the existence, and nature, of early life." Schopf, J. W. (1992:39, 40).
A reconstruction of the 3,500 million years old environmental setting of the Apex chert displayed at the U.S. Natural History Museum, Washington, D.C., and designed by K. M. Towe of the Smithsonian Institution. From J. W. Schopf (1999:91) Fig. 3.11. In the foreground: The round, cake-like structures in the low water at the shore are Precambrian stromatolites. There, photosynthetic one-celled organisms and other kinds of bacteria are living. They have not changed at all. They do not know anything about evolution, about evolving upward into higher forms of life. In the background, active volcanoes spouting their gases into the air.
Where has one found now the oldest microfossils? What do they show us?
Prof. J. Schopf: "The best evidence - evidence that seems fully convincing - comes from microfossils. In particular, narrow, microscopic bacterium- or cyanobacterium-like filaments have been found petrified in sediments of both the Swaziland Supergroup (in finely layered black cherts of the Onderwacht Group), and the Pilbara Supergroup (in gray to black carbonaceous cherts of the Warrawoona Group).
"Isotopic studies indicate that organic matter in both deposits, the minute petrified filaments, like fossils that have been preserved in a similar manner in younger sediments (petrified woods, for example), are composed of carbonaceous, organic material. Some of the Swaziland filaments appear to be surrounded by a thin, hollow, organic tube, an organization unlike that occurring in minerals, but one that is well known in modern filamentous bacteria and cyanobacteria (in which strands of cells are encompassed by an originally umcilagelike, tubular, organic 'sheath'). And the Pilbara filaments are composed unquestionably of distinct, organic-walled cells (Figure 2.5 in his book), arranged in single-file rows like beads on a string - definitely a nonmineralic organization, but one that is characteristic of the great majority of living, filamentous microbes. Furthermore, the particular arrangement of these cells, and the occurrence of partially divided pairs of cells in some of the Pilbara filaments, indicates that they were produced by the same type of cell division as that occurring in living prokaryotic microorganisms.
"Six different types of fossil filaments have been discovered in the Pilbara cherts, classified by the size and shape of their cells, including their end cells, which in some filament types are rounded or conical (Figure ee2.5A). Interestingly, meticulous comparison of these filaments with microbes living today show that the majority of these fossils are similar in cellular detail to particular species of living cyanobacteria. (cyano = greenish-blue).
"Moreover, the occurrence of stromatolites in units both of the Swaziland and Pilbara Supergroups, and the carbon isotopic signature of the organic matter preserved in both of these sequences, are also consistent with the possible presence of cyanobacteria (although they do not prove that cyanobacteria were certainly present, since both the stromatolites and the carbon isotopic values might otherwise have been produced by photosynthetic bacteria). Thus the evidence suggests that cyanobacteria may have existed as early as about 3 500 Ma ago. And this is an intriguing possibility, because if these fossil microbes were, in fact, cyanobacteria, they would have to have been capable of oxygen-producing photosynthesis - their presence would indicate that microorganisms with this advanced biochemical capability had already evolved by this early stage in the history of life.
"Considering their very great geologic age, the Swaziland and Pilbara filaments, the oldest fossils now known, seem surprisingly advanced. Some are virtually indistinguishable in form from microbes living today and, apparently, their cells divide by the same processes, they lived in the same types of environments, and they carried on the same sorts of metabolism as those of their modern microbial look-alikes.
"Evidently, shallow-water Archean seas were inhabited by complex, biologically diverse communities of stromalite-forming microorganisms, self-contained ecosystems that included both photoautotrophic producers and anaerobic, heterotrophic consumers (microbes that recycled the photosynthetically produced foodstuffs), and that possibly even included advanced oxygen-producing cyanobacteria." Schopf, J. W. (1992:42-44).
Since when have there been cyanobacteria on earth, making oxygen? What do the banded iron-formations of the Early Precambrian show us?
Prof. J. W. Schopf: "Evidence from the highly metamorphosed Isua rocks indicates that both water and carbon dioxide were present in the environment at least as early as 3750 Ma ago. Because the starting material required for oxygenic photosynthesis were already present at that remote time (and because sunlight, needed to power photosynthesis, has existed since the earlier time of formation of the solar system), it is worth asking whether oxygen-producing cyanobacteria might also have existed in Isual time. In fact, this is certainly conceivable, a possibility suggested by the occurrence of oxidized iron minerals, occurring in banded iron-formations (BIFs) (Fig. 2.2), in the Isua sequence.
"What are banded iron-formations (abbreviated BIFs), and how do they enter the picture? ... Typically, the banding is produced by an alternation of iron-rich and iron-poor layers in the rock, and because the iron-rich layers are composed of fine rustlike particles of iron-oxides (particularly the iron-rich hematite, Fe2O3; and, in some deposits, the mineral magnetite, Fe3O4), the iron-rich layers have a distinctive dull to bright red color.
"The iron minerals are formed when iron, produced by volcanic activity and dissolved in oceanic waters, combines with molecular oxygen, a chemical reaction that normally occurs in the upper reaches of the water column, where oxygen is present. Because the resulting iron oxides are highly insoluble in seawater, a fine rain of minute rusty particles falls onto the ocean floor." (1992:44).
From where has this iron in the banded iron-formations come? And from where has this oxygen come, which then oxidized this iron?
Prof. J. W. Schopf: "For a great number of reasons, it is well established that the predominant source of atmospheric O2 is oxygen-producing photosynthesis. From this it follows that prior to the origin of oxygenic, cyanobacterial photosynthesis, the atmosphere must have been virtually devoid of free oxygen; the planetary environment must have been anaerobic (Fig. 2.2); and the oxygen required for the deposition of BIFs must have been in very short supply.
"Hence, if BIFs were widespread during Isual time, and if these iron oxide-rich units were deposited by the same mechanism as that occurring later in the Precambrian, then their occurrence would seem to require the presence of oxygen-producing photosynthesizers 3750 Ma ago." (1992:45, 46).
The cyanobacterium makes free oxygen. If it has been there already some 3.5-3.8 billion years ago, why, then, was there so little free oxygen in the atmosphere of the Early Precambrian? And how does the cyanobacterium "make" this free oxygen?
Prof. J. W. Schopf: "In addition to nitrogen, water vapor, and carbon dioxide, major constituents of the atmosphere, volcanoes give off smaller amounts of other gases. Certain of these, such as hydrogen, methane, carbon monoxide, and hydrogen sulfide, can readily combine with molecular oxygen. Thus, for example, hydrogen (H2) combines with oxygen, that is, it is oxidized, to yield water (H2O); both methane (CH4) and carbon monoxide (CO) are oxidized to produce carbon dioxide (CO2); and hydrogen sulfide (H2S) is oxidized to yield soluble sulfate (SO42-). Such volcanic gasses, therefore, are effective oxygen scavengers, serving as one of the three oxygen sinks in the early environment.
"Oxygen produced by photosynthesis was also scavenged biologically. During oxygenic photosynthesis, light energy is used to split water (H2O) into hydrogen and oxygen; the hydrogen from the water is combined with carbon dioxide to produce organic matter (usually written in shorthand as sugar, 'CH2O'); and the oxygen from the water is released into the environment, an unused by-product of the photosynthetic process. Thus, ... the net chemical reaction of the cyanobacterial photosynthesis is
(light energy) + water carbon dioxide > organic matter +oxygen.
Aerobic Respiration. What happens during aerobic respiration, when the organisms breathe?
Prof. J. W. Schopf: "This is the exact opposite of the biochemical reaction that takes place during aerobic respiration. When organisms 'breathe' (a capability of all aerobic organisms, whether they are facultative or obligate aerobes, and even including plants, cyanobacteria, and aerobic bacteria), they oxidize organic matter to release energy by carrying out this net chemical reaction:
Organic matter + oxygen > water + carbon dioxide + (cellular energy).
Thus, a second oxygen sink was provided by microorganisms capable of carrying out aerobic respiration. The earliest such microbes were no doubt facultative, respiring aerobically, when oxygen was available, but switching to anaerobic fermentation, when local oxygen levels became low (for example, during times of active volcanism, when the oxygen was scavenged by volcanic gasses). Facultative aerobes, effective oxygen scavengers, sponged up the photosynthetically produced oxygen before it could build up in the atmosphere." (1992:47, 48).
Dissolved Iron. How does the dissolved iron swallow up free oxygen molecules?
Prof. J. W. Schopf: "The third of the three oxygen sinks was provided by the dissolved iron that had accumulated in the world's ocean basins throughout early earth history. As the iron washed up into areas where photosynthetically produced oxygen was available, the oxygen combined with the iron to form insoluble iron oxide minerals; the mineral particles settled to the ocean floor as a fine, rusty rain; and the BIFs [Banded Iron Formations] were deposited. In effect, the photosynthetically produced oxygen was scavenged from the oceans and buried forever in the form of rust." (1992:48).
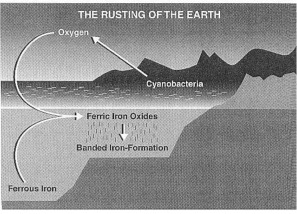
The Precambrian Earth began to rust, when iron in the ocean began to oxidize, when it combined with oxygen, released by cyanobacteria. These ferric iron oxides then sank down to the bottom forming the banded iron formations (BIF). Without them, our Present civilization, based on iron and steel, could never have arisen. From J. W. Schopf (1999:172) Fig. 6.2
Has the prokaryotic cell evolved during the nearly two billion years of Proterozoic time? In other words: Have today's bacteria come from a simple, primitive ancestral cell? And has this first cell of the Early Precambrian evolved then into plants, animals, and human beings?
Prof. J. W. Schopf: "The answer is: A lot can be said: even at present, a great deal is known. Surprisingly, however - in fact, absolutely remarkably - cyanobacteria in particular, and perhaps all prokaryotes in general, seem to have evolved hardly at all between early in the Proterozoic and the present day! That, indeed, is strange. ... Judging from their morphology, living and Proterozoic cyanobacteria are virtually indistinguishable. Figure 2.8 (in his book) shows just four of a large number of examples that might be cited illustrating this uncanny similarity between the modern and the fossil.
"Compare, for example, the living cyanobacterium Lynbya, shown in Figure 2.8A, with fossil Paleolyngbya, shown in Figure 2.8B: Both are about the same size; both are composed of single-file rows of disk-shaped cells; both have rounded terminal cells; both are enclosed by tubular organic sheaths. The fossil filaments (Figure 2.8B) are nearly one billion years old, but if they were alive today, they surely would be placed in the same genus, and perhaps even the same species, as the modern filaments.
"Or compare the living coiled cyanobacterium Spirulina (Figure 2.8C) with its 850-Ma-old fossil look-alike (Figure 2.8D); or the sheath-enclosed, four-celled colony of modern Gloeocapsa shown in Figure 2.8E, with the notably similar 1550-Ma-old Gloeocapsa-like colony shown in Figure 2.8F; or compare the many-celled colony of living Entophysalis (Figure 2.8G) with its fossil counterpart, the 2150-Ma-old colonial Eoentophysalis (Figure 2.8H).
"Other evidence is also available. The range of diameter and pattern of size distribution of the tubular sheaths of living cyanobacteria and of comparable fossil prokaryotic filaments are essentially identical (Figure 2.9). Moreover, there is no discernable trend, no evident pattern of evolutionary change, in the size either of fossil tubular sheaths or of fossil cellular filaments over all of Proterozoic time, an eon some four times longer than the entire Phanerozoic!" (1992:50, 52).
What do you conclude from this?
Prof. J. W. Schopf: "The conclusion seems inescapable - whether the organisms are filamentous or spheroidal, whether they are straight or coiled, whether they occur in few- or in many-celled colonies, and regardless of cell size or cell shape or the nature of their extracellular sheaths - the morphology of cyanobacteria has changed little, if at all, since Proterozoic time."
"But the size, shape, and cellular organization of cyanobacteria have remained unchanged over periods of more than one, and in some cases more than two billion years. ... In addition to morphology, there are many other obvious similarities between living and Proterozoic cyanbacteria (for example, the environments in which they occur; their ability to form stromatolites; their concentration at the growth surface in microbial communities, and the particular types of cyanobacteria, the families and genera, occurring in such communities). Hence, all things considered, it seems quite likely that the fundamental nature of such prokaryotes has evolved little, if at all, since early in Proterozoic time." (1992:53).
What do you conclude now from your findings? Since when has there been life on earth?
J. W. Schopf and C. Klein: "The cell sizes of the majority of fossil trichomes seem more suggestive of cyanobacterial, than of bacterial, affinity. ... Therefore, by analogy with extant microorganisms, the majority of the filamentous morphotypes now known from the Warrawoona cherts seem more likely to be of cyanaobacterial, than of bacterial, affinities. Although some extant cyanobacteria are capable of facultative anoxygenic photosynthesis, the ability to carry out oxygen-producing photoautotrophy is a universal characteristic of this group. Based on these observations, it thus seems a plausible inference that cyanobacteria, and therefore oxygen-producing photosynthesizers, are likely to have been represented in the Warrawoona biota and, hence, that this relatively advanced physiological evolution may have been attained at least as early as ~3500 Ma ago.
"It is evident that as early as ~3500 Ma ago microbial communities were extant, morphologically varied, and possibly physiologically advanced, and that Archean microfossils, both filamentous and colonial, are notably similar in morphological detail to extant prokaryotes, a similarity evidently extending even to the mechanisms of cell division." Schopf and Klein (1992:38, 39).
"The known stromatolitic, microbial, and carbon-isotopic fossil records seem to establish firmly the existence of photoautotrophic prokaryotes (viz., photosynthetic bacteria and, possibly, O2-producing cyanobacteria) as early as 3.5 to 3.4 Ga ago. ... Anaerobic bacteria are inferred to have been extant as early as ~ 3.5 Ga ago. Indeed, the occurrence of reduced carbon in the highly metamorphosed, ~3.8 Ga-old Isua Supracrustal Group of south-western Greenland (Schidlowski et al. 1983) - if not a product of abiotic processes - may reflect the presence of such microbes even earlier in geologic time."
"The occurrence of Archean stromatolites, known from earliest sediments of the ~3,5 -Ga-old Warrawoona Group of Western Australia, is thus strong evidence for the presence of photosynthetic bacteria (and is equally consistent with the existence of ... O2-producing cyanobacteria). Similarly, at least the narrower filamentous microfossils in some Early Archean stromatolite-like laminae (e.g, those in cherty sediments of the ~3.4 Ga-old Ondervacht Group of South Africa) are morphologically similar to extant photosynthetic bacteria (viz., Chloroflexus). And carbon-isotopic ratios in kerogens from both Warrawoona and Onverwacht Group sediments are consistent with photoautotrophic carbon fixation (e.g., by photosynthetic bacteria and/or cyanobacteria).
"Thus, the conclusion seems clear: anoxygenic photoautotrophic bacteria had become established at least as early as ~3.5 Ga ago. Moreover, as suggested by the occurrence of graphitic carbonaceous matter in ~3.8 Ga-old sediments of the Isua Supracrustal Group, such organisms may have been extant even earlier. ... The paleoenvironmental, stromatolitic, microfossil, and carbon-isotopic data now available from units 3.5 to 3.3 Ga in age (as well as the occurrence of banded iron-formations and reduced carbon in these and sediments as old as ~3.8 Ga) are all consistent with the Early Archean existence of oxygen-producing cyanobacteria. ... Cyanobacteria are inferred to have been extant during the Late Archean with their possible presence extending well into the Early Archean." Schopf and Klein (1992:587-590).
![]()
Domical and columnar stromatolites built today at Shark Bay, Western Australia (A), look almost identical to fossil examples billions of years old (B, from the 2,300-million-year-old Transvaal Dolomite of Cape Province, South Africa. The geological hammer in (A) at right center, shows the scale of both photos. From J. W. Schopf (1999) Fig. 7.7.
Many of the nearly 300 prokaryotic taxa of Proterozoic microbial species one may compare in morphological detail to specific, modern microorganisms. Many of the fossil cyanobacterial species, which one now knows, seem to have lived during large parts of Proterozoic time ranges (Schopf and Klein 1992:596). Does this mean then that they are also biochemically similar?
Schopf and Klein: "With regard to genetics, little can be said aside from the observations that (i) morphologically close comparability between modern and fossil taxa seems likely to reflect fundamental similarities in the genetic and developmental controls governing their morphology; (ii) direct fossil evidence seems to indicate that the modern mechanisms of prokaryotic (including cyanobacterial) cell division were already in place as early as 3.4 Ga ago; and (iii) virtually the entire range of morphological products of such cell division exhibited among modern cyanobacteria is also exhibited among Proterozoic taxa, whether considered in terms of the shape of the medial (e.g., discoidal, quadrate, barrel-shaped, elongate) or terminal cells (globose, conical, rounded, blunt-ended) of cyanobacterial trichomes, of the cell morphologies in non-filamentous colonies (coccoid or ellipsoid), or of the organization of cells that make up cyanobacterial colonial aggregates (e.g., occurring in unseriate, unbranched trichomes; irregular colonies; and in tabular, decussate, cuboidal, rosette-like, and similar highly ordered colonies).
"Data from the fossil record are consistent with the proposition that these characters have not changed significantly over geologic time; no current evidence refutes (or even seriously challenges) the correctness of this proposition... Several lines of evidence suggest, however, that it is unlikely that either the biochemistry or the physiology of cyanobacteria has changed appreciably since at least ~2.1 Ga ago, the beginning of the relatively well-documented fossil record that links cyanobacterial fossils into an evolutionary continuum merging with the present." Schopf and Klein (1992:596, 597).
Has this "evolutionary continuum" caused the first living cell to evolve into higher forms of life, into plants, animals, and human beings?
J. W. Schopf and C. Klein: "With respect to biochemistry, for example, the range of morphologies exhibited by Proterozoic cyanobacterial sheaths (viz., from thin, diffuse, and diaphanous to thick, robust, and multilamellated) essentially identical to that characteristic of the sheaths enclosing extant coccoid and filamentous cyanobacteria, is consistent with an original polysaccharide composition like that of the modern analogues, as is the commonly reported preferential preservation of such sheaths both in the Proterozoic fossil record and at depth in modern microbial mats.
"In terms of physiology, the occurrence of the isoprenoid hydrocarbons pristane and phytane (in part presumably chlorophyll-derived) in fossil cyanobacterium-containing stromatolitic Proterozoic sediments, the carbon-isotopic signatures of kerogens isolated from such units, and the laminar, mat-forming orientation of stromatolitic cyanobacterial photosynthesis, the oxygen thus liberated presumably being evidenced by the occurrence of oxidized Proterozoic sediments such as banded iron-formations and red beds.
"Similarly, where the paleoecology/paleobathymetry of fossiliferous units can be inferred confidently, all Proterozoic sediments containing non-transported fossil cyanobacteria appear to have been deposited within the photic zone, the fossils occurring, like modern cyanobacteria, as components of stromatolitic biocoenoses. Moreover, whether at the familial or generic level, the compositions of Proterozoic and modern mat-building cyanobacterial communities are notably similar, both being dominated by oscillaltoriaceans (e.g., Oscillatoria, Lyngbya, Phormidium, Microcoleus) with subordinate chroococcaceans (e.g., Microsystis, Aphanocapsa, Gloeocapsa, Chroococcus) and, in some settings, entophysalidaceans (e.g., Pleurocapsa).
"Finally, the sparse evidence now available from extract organic geochemistry (e.g., Proterozoic distribution of hopanes, steranes, and other biomarkers)... Although not definite, is also consistent with the postulated absence of significant biochemical or physiological cyanobacterial evolution.
"In short, therefore, and despite the fact that indirect evidence and 'consistency argument' like the foregoing are less than compelling, it seems a reasonable supposition that the hypobradyletic (= little or no) evolution of Proterozoic cyanobacteria was characteristic not only of their morphology, but of at least the most basic aspects of their genetics, biochemistry, and physiology as well." Schopf and Klein (1992:596, 597).
Since when have there been cyanobacteria on earth, formerly known as blue-green algae? Where have they lived? And how complex were they?
J. William Schopf, at the Molecular Institute, University of California, Los Angeles, reports in Science, 30 April 1993, p. 640-646 about his new findings in northwestern Australia:
"Eleven taxa (including eight heretofore undescribed species) of cellularly preserved filamentous microbes, among the oldest fossils known, have been discovered in a bedded chert unit of the Early Archean Apex Basalt of northwestern Western Australia. This prokaryotic assemblage establishes that trichomic cyanobacterium-like microorganisms were extant and morphologically diverse at least as early as ~3465 million years ago and suggests that oxygen-producing photoautotrophy must have already evolved by this early stage in biotic history.
"The most promising terrain for such studies is that of the Pilbara Block of Northwestern Western Australia, a region underlain by a 30-km-thick sequence of relatively well-preserved sedimentary and volcanic rocks that are ~3000 to 3500 million years old. From this region, I describe a diverse assemblage of filamentous microbial fossils detected in the Early Archean (~3465 million years old) Apex chert, cellular prokaryotes more than 1300 million years older than any comparable suite of fossils previously reported from the geological record.
"This suggests that cyanobacterial oxygen-producing photosynthesizers may have already been extant this early in Earth history. ... Sheath-enclosed colonial unicells occurring in ~3465-million-year-old sedimentary rocks of the Towers Formation, also of Western Australia; and narrow nonseptate bacterium-like filaments from ~3450-million-year-old units of the Swaziland Supergroup of South Africa.
"A maximum age for the Apex chert of ~3470 Ma is constrained by U-Pb zirkon ages... A minimum age for the fossiliferous rocks of ~3460 Ma is provided by a U-Pb-zirkon date of 3458±1.9 Ma for the immediately overlying Panorama Formation. ... Thus, the age of the fossiliferous Apex chert is evidently about 3465 Ma." (Schopf, J. W. (1993:640, 641).
Why can be sure that they really are the remains of bacterial cells? What scientific proof is there?
Prof. J. W. Schopf: "As discussed below, their evident cellular organization, and their morphological complexity and similarity to younger prokaryotes, both fossil and modern, firmly established their biogenicity. ... Whether the microfossils are greatly older than or essentially just as old as the Apex chert is not known." (1993:641, 642).
"Eleven taxa of filamentous, dark brown to black carbonaceous microfossils, including eight new species, have been identified in the deposit. Solitary unicell-like spheroids of possible but uncertain biological origin also occur.
"Single taxa (especially, Primaevifilum minutum, n. sp.; P. laticellulosum, n. sp. and P. attenuatum n. sp.) or particular pairs or groups of taxa (for example, P. delicatulum and Archaeoscillatoriopsis disciformis, n. gen. N. sp.; or P. delicatulum, P. amoenum and P. conicoterminatum) tend to predominate in individual clasts. Although possibly representing a benthic microbial community that was loosely organized and embedded in mucilage, the filaments exhibit neither the subparallel orientation nor the laminar organization typical of most stromatolitic microbiotas. Microfossils have not been detected in stromatolite-like laminated clasts that also occur in the unit. ... The incomplete preservation of the Apex fossils suggests that the original assemblage probably included more taxa than the 11 species identified.
"As currently documented, the fossil record is more or less continuous and relatively well known from about 2100 Ma to the present, beginning with the diverse microbiotas of the ~2100-million-year-Belcher Group and the ~2080-million-year-old Gunflint Iron Formation, both of Canada. But the fossil record from the greater than 1300 million years intervening between these deposits and the Apex chert is essentially undeciphered. Although there is thus a profound gap in the record, the morphological similarity of the Apex fossils to septate filamentous prokaryotes, both Proterozoic and modern, indicates that they are almost certainly prokaryotes and part of an evolutionary continuum that extends from the Early Archaean to the present. This interpretation seems supported by the occurrence in Apex filaments of bifurcated cells and cell pairs, that evidently reflect the original presence of partial septation and, thus, of cell division like that occurring in extant prokaryotic filaments." (1993:642, 643).
How do these bacterial cells, about 3.5 billion years old, compare with modern bacterial cells?
Prof. J. W. Schopf: "In comparison with modern prokaryotes, most of the Apex microbes particularly resemble trichomic (nonensheathed or thinly ensheathed) oscillatoriacean cyanobacteria. Cell widths of the Apex taxa range from 0.5 µm to 19.5 µm and average ~5.0 µm (Table 1). Modern filamentous bacteria tend to be quite narrow, predominantly < 1.5 µm in diameter, whereas most oscillatoriacen trichomes are notably broader. On the basis of morphometric analyses of more than 500 taxa of modern filamentous microbes, I have suggested that fossil septate filaments < 1.5 µm wide be regarded as 'probable bacteria,' those 1.5 µm to 3.5 µm wide as (undifferentiated) 'prokaryotes,' and those > 3.5 µm broad as 'probable cyanobacteria'.
"Applying these criteria to the Apex fossils, I interpret two taxa (Archaeotrichion septatum, n. sp., and Eoleptonema apex, n. sp.) as probable bacteria; two taxa (Primaeevifilum minutum, n. sp., and P. delicatum) as either bacteria or cyanobacteria; and the remaining seven species, nearly two thirds of the taxa (and 63 percent of measured specimens) as probable cyanobacteria.
"Because of the size ranges of filamentous bacteria and cyanobacteria overlap, the suggested affinities are not absolute. Nevertheless, the pattern of size distribution exhibited by the Apex assemblage is more like that of modern oscillatoriaceans than of noncyanobacterial prokaryotes. Furthermore, several of the Apex taxa, particularly those with broad trichomes (Primaevifilum laticellulosum, n. sp.; Archaeoscillatoriopsis grandis, n. gen., n. sp.; and A. maxima, n. gen. N. sp.) differ in cell size from almost all bacteria but are essentially indistinguishable from specific oscillatoriaceans, both Proterozoic (Oscillatoriopsis spp.) and modern (Oscillatorioa spp.). If the Apex filaments had been discovered in later Precambrian sediments, in which fossil oscillatoriaceans are well known and relatively widespread, or if they had been detected in a modern microbial community and morphology were the only criterion by which to infer biological relationships, the majority would be interpreted as oscillatoriacean cyanobacteria." Schopf, J. W. (1993:643).
"The range of morphologies exhibited by the Apex filaments indicates that if the majority are oscillatoriaceans, this primitive family of filamentous cyanobacteria are capable of temporarily carrying out anoxic (bacterial) photosynthesis, oxygen-producing photoautotrophy is a universal, presumably self-evolving characteristic of this group. The presence of diverse oscillatoriaceans in the Apex assemblage would thus seem to imply that this relatively advanced level of physiological evolution had been attained at least as early as ~3465 Ma." (1993:643, 644).
What proves that there have been oxygen-producing oscillatoriaceans (= cyanobacteria) so early in earth's history, some 3 500 million years ago?
Prof. J. W. Schopf: "Four other lines of evidence seem consistent with the possible Early Archean existence of O2-producing oscillatoriaceans: (i) Early Archean stromatolites were presumably produced by photoautotroph-dominated microbial communities. (ii) The reactants required for oxygenic photosynthesis, CO2 and H2O, and materials possibly representing products of this process, sedimentary organic matter and oxidized iron minerals, were present in the Early Archean environment. (iii) The isotopic composition of Early Archean organic and carbonate carbon are evidently indicative of photosynthetic CO2-fixation like that occurring at relatively high CO2 concentrations in extant microbial populations. (iv) Calculations based on models of the early global ecosystem, and cerium and europium concentrations in Archean banded iron-formations suggest that O2-producing photosynthesis and aerobic respiration both date from the Early Archean. These additional lines of evidence, however, are not conclusive; all but the latter, which necessarily incorporates model-dependent uncertainties, would be equally consistent with the presence of solely anoxic bacterial photosynthesizers." (1993:644).
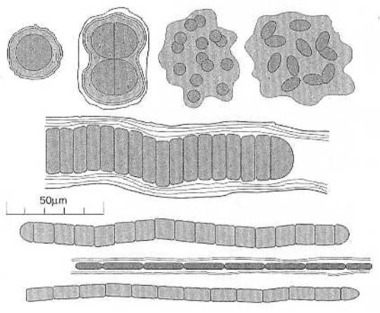
Cyanobacteria, coccoidal (ball-shape), ellipsoidal, and filamentous (thread-shape). Cyanobacteria come in a variety of shapes and sizes and are often surrounded by or embedded in secreted layers of sticky mucilage (slime). Each slice in the thread- or sausage-like forms is a photosynthetic cyanobacterium. The cyanobacteria changes the energy of the sunlight into organic substances. And it produces free oxygen. From J. W. Schopf (1999:187) Fig. 7.1. The first cyanobacterium, some 3.7 billion years ago, has been just as complex as those still living today. It has not evolved at all into any higher forms of life, because it was perfectly adapted to its environment since it began.
Also the fossilized remains of different species of the cyanobacterium, the greenish-blue bacterium have been found in Northwestern Australia. The cyanobacterium Oscillatoria, which one has found at Warrawoona, N.W. Australia, is about 3.5-3.5 billion years old. Professor J. W. Schopf (1983, 1992) has reported about these finds. How large are these oxygen-producing cyanobacteria? How long is their genome, their genetic code? And how much information does it contain? In other words: What was needed, to put their DNA-pairs into the right order? Let us look briefly at a few of them:
Primaefilum conicoterminatum, 4.5 to 7.0 µm wide and 3.0 to 4.5 µm long. Average 5.9 µm wide and 3.9 µm long. Schopf and Klein (1992:35). Modern analogue: cyanobacterium Oscillatoria acuta.
Primaevifilum laticellulosum, 6.0 to 8.5 µm wide, 2.5 to 5.0 µm long, average 7.0 µm wide, 3.5 µm long. From Warrawoona, Apex Chert, 3.5 billion years old. Modern analogue: cyanobacterium Oscillatoria tenus. Schopf, J. W. (1993:641, 645).
Genus Archaeoscillatoriopsis, 3.0 to 19.5 µm wide, 0.8 to 6.0 µm long. Modern analogue: Oscillatoria ssp. oscillatoriaceans. Warrawoona, Apex Chert, 3.5 billion years old. Schopf, J. W. (1993:641, 645).
Archaeoscillatoriopsis disciformis, 3.0 to 5.5 µm wide, 0.8 to 2.2 µm long, average 4.2 µm wide, 1.2 µm long. Modern analogue: modern cyanobacterium Oscillatoria grunowiana. From Warrawoona, Apex Chert, 3.5 billion years old. Schopf, J. W. (1993:641, 645).
Archaeoscillatoriposis grandis, 8.0 to 11.5 µm wide, and 1.0 to 3.5 µm wide, average 9.0 µm wide, 2.0 µm long. Modern analogue: modern cyanobacterium Oscillatoria chalybea. Schopf, J. W. (1993:641, 645). From Warrawoona, Apex Chert, 3.5 billion years old.
Archaeosillatoriopsis maxima, 15.0 to 19.5 µm wide and 3.0 to 6.0 µm long, average 16.5 µm wide and 4.5 µm long. Modern analogue: modern cyanobacterium Oscillatoria antillarum. From Warrawoona, Apex Chert, 3.5 billion years old. Schopf, J. W. (1993:641, 645).
How long is the genome of this oxygen producing cyanobacterium? And how much information does it contain? - Michael Herdman and co-workers (1979:80) have determined the genome size of six strains of Oscillatoria. It ranges from 2.50·109 to 4.38·109 dalton. Its average size is 3.62·109 dalton. 3.62·109 Da : 660 Da/bp = 5 484 848 bp log 4 = 103 303 207 bit.
This means: Already 3.5 billion years ago the earth had cooled down enough. There were many different genera and species of the cyanobacterium Oscillatoria. Many of these species are still living today. They have not changed at all, because they have been perfectly adapted to their environment, from the beginning! Their genome had an average size of 5 484 848 base pairs. Most of the cell's genetic information is stored in its genome. It contains at least 103 303 207 bit of information or yes/no decisions!
Synechocystis. Also the oxygen producing cyanobacterium Synochocystis aquatilis one has found in the 3.3-3.5 billion year old sediments at Warrawoona, N.W. Australia. It has a diameter of 5.0 to 6.0 µm. Schopf, J. W. (1983) Table 9.2. - How long is its genome? And how much information does it contain?
Synechocystis, with a diameter of 6-7 µm (PCC 6806) has now a genome size of 2.31·109 Da. A cell with a diameter of 5.0-6.0 µm would have then a genome size of about 2·109 Da. Herdman, M. et al. (1979:67). 2·109 Da : 660 Da/bp = 3 030 303 bp log 4 = 101 824 424 bit.
Synechocystis has now a genome size of 1.79·109 to 3.5·109 Da. Its mean size is 2.29·109 Da. 2.29·109 Da : 660 Da/bp = 2 712 121 bp log 4 = 102 088 965 bit.
Result
Why is there life on earth? Why are there one-celled organisms? What do the findings at the Warrawoona Group in Northwestern Australia show us, which are said to be 3.3-3.5 billion years old? Do they prove that life on earth has evolved from the first cell, from a common ancestor?
The fossil evidence at Warrawoona clearly disproves the beliefs of the evolutionists. There is no proof whatsoever in the sediments of the Early Precambrian time, some 3.3-3.5 billion years ago, that all life on earth has come from a common ancestor, from the "first cell". To the contrary. Right from the start there are many different genera and species of bacteria and archaea, like today.
There is no proof whatsoever in the world's oldest sediments that inorganic matter organized itself from the not-living to the living, from the lower to the higher, and from the simple to the complex. To the contrary. Only remains of perfect one-celled organisms, of bacteria and archaea have been found. They are identical with many of the genera and species, which are still living today. They do not know anything about evolution. They have remained, what they were, right from the beginning. Also the banded iron formations (BIFs) and the organic carbon at Warrawoona do prove that these "first" cells were already as complex, as the cells, which are living now, some 3.3-3.8 billion years later.
In the sediments of the Early Precambrian there is no transition from the not-living to the living, from the simplest atoms and molecules to man. There are only perfect one-celled organisms, perfectly adapted to their environment from the start. The first living cell on earth has been already more complex, than anything, which man has been able to make till now. The findings near the beginning of the world clearly prove that God has thought out and made them.
What have other scientists found out about life on earth, and how it has arisen?
Cyanobacteria from Early Proterozoic time, some 2000 to 2500 million years ago, after Archean time: How complex were these one-celled organisms? How much have they changed since then? Have they, perhaps, evolved into higher forms of life?
Andrew H. Knoll, Botanical Museum, Harvard University, Cambridge, MA, USA, and Stejepko Golubic, Dept. of Biology, Boston University, Boston, MA, USA, report about "Proterozoic and Living Cyanobacteria.":
"Essentially all of the salient morphological features used in the taxonomic classification of living cyanobacteria can be observed in well-preserved microfossils, despite the pervasive influence of taphonomic (= when they died and were buried) alteration of Proterozoic microbial remains. ... Some blue-greens (= cyanobacteria) form extra-cellular sheaths or envelopes that are relatively resistant to post mortem alteration. These remains may be preserved three dimensionally, providing detailed information about cell size, shape, and filament of colony form." (1992:453).
A. H. Knoll and S. Golubic: "One of the first, and still one of the most illuminating comparisons of a Proterozoic microfossil population and a specific recent cyanobacterium is that between the fossil genus Eoentophysalis and its modern counterpart, Entophysalis. ... The genus Entophysalis is characterized by its coccoidal cells that produce by binary fission in three planes. External envelopes are persistent through several cell divisions, giving rise to cell packets that preserve a record of their divisional history.
"Modern Entophysalis species occur in a variety of peritidal and playa lake environments, but they are particularly abundant and conspicuous along the margin of water bodies characterized by elevated salinity. In such environments, Entophysalis populations can form structurally competent and sedimentologically distinctive mats, and participate in the accretion and lithification of stromatolitic structures." (1992:455).
"Entophysalis, then, is a cyanobacterium that is well characterized morphologically, ecologically, and taphonomically; significantly, it is also a cyanobacterium whose morphology is diagnostic for the phylum (i.e., the distinctive colony form of Entophysalis is not known to be matched in other eubacterial phyla). It is, therefore, an ideal focus for comparison with fossil populations, and research published over the last 15 years indicates that close fossil counterparts to Entophysalis are common in Proterozoic strata.
"The genus Eoentophysalis was established for large populations in the Early Proterozoic Belcher Supergroup, Canada (Hofmann 1976). Belcher Eoentophysalis populations conform to living Entophysalis species in the details of their cell size and shape, division pattern and life cycle, envelope characters and colony form, taphonomic pattern, and inferred ecology and environmental distribution (Golobic and Hofmann 1976), leaving little doubt about the systematic paleontology and physiological attributes of the nearly 2000-Ma-old fossils. Further Eoentophysalis populations, representing several species, have been described from the Middle Proterzoic McArthur (Muir 1976), Nathan (Oehler 1978), and Dismal Lake Groups (Horodysk and Donaldson 1983), and Gaoyuzhuan Formation (Zhang 1981). Late Proterozoic examples are known from the Narssarssuk (Strother et al. 1983) and Bitter Springs formation (Knoll and Golubic 1979). ... In all cases, the fossil populations conform morphologically, ecologically, and taphonomically to their living counterpart." Knoll, A. H. and S. Golubic (1992:455).
Knoll and Golubic: "Morphologically distinctive microfossils assigned to the genus Polybessurus were first described from silicified Upper Proterozoic carbonates of South Australia by Fairchild (1975). More recently, Green et al. (1978) have published detailed observations and interpretations of large populations from the Upper Proterozoic Eleonore Bay Group, East Greenland. As in the case of Eoentophysalis, Polybessurus populations can be matched with living cyanobacteria whose morphology is diagnostic for the phylum (Green et al. 1987).
"Polybessurus individuals were spheroidal unicells whose systematic envelope production resulted in the formation of gelatinous stalks that elevated the cell above the sediment interface. Reproduction is inferred to have been by baeocyte formation. Polybesssurus fossils sometimes occur as isolated individuals within microbial mats, but more conspicuously, large populations formed crusts on fine grained carbonate substrates in peritidal environments. To date, populations that fit this description are known from Middle and Upper Proterozoic formations in East Greenland (Green et al. 1987), Svalbard (Butterfly and Knoll, unpubl. data), Australia (Fairchild 1975), and Arctic Canada (Butterfield et al., unpubl. data).
"Physically analogous environments on the present-day Bahama Banks host cyanobacteria that, except for their smaller size, are comparable to Proterozoic Polybessurus populations (Green et al. 1987). This modern counterpart is as yet undescribed, but is similar to the genus Cyanostylon. It forms gelatinous stalks by asymmetric gel excretion, as sequentially produced envelopes rupture at their upper, thinner side to leave behind the stack of inverted gelatinous cones. The presence of these living cyanobacteria in environments that could be predicted on the basis of the Proterozoic fossil record once again leaves little doubt about the systematic and ecological interpretation of the fossils." Knoll and Golubic (1992:455, 456).
Knoll and Golubic: "Brief mention of a third compelling ancient/modern comparison should serve to cement the point. Ooids found at present in shallow subtidal environments of the Bahama Banks are riddled by endolithic cyanobacteria; up to six species may occur within a single coid. These cyanobacteria are morphologically, developmentally, and behaviorally distinctive. Late proterozoic ooids from the Eleonore Bay Group, East Greenland (Green et al. 1988), and the Backlundtoppen Formation, Svalbard (Knoll et al. 1989), host abundant and diverse endolithic microfossils whose morphology, development and reproduction, orientation within the substrate (from which behavior can be inferred), and environmental distribution compare closely with those of the populations found in Recent ooids.
"Several species of the cyanobacterial genus Hyella occur in Bahamian endolithic communities. These populations are united by several characteristics: the presence of pseudofilaments composed of spheroidal to cylindrical coccoidal cells, pseudofilament branching by cell slippage or apical cell dichotomy, the presence of degradation-resistant envelopes that define cell shapes and positions, binary fission accompanied by baeocyte formation, and a distinctive negative phototropism and attendant polarity reversal in development.
"Four species attributable to the genus Eohyella occur in Late Proterozoic ooids from Svalbard and East Greenland. Two prominent populations are closely comparable to Hyella gigas and H. caespitosa, while a third has a living counterpart in an as yet unnamed Hyella species. This comparison across 700-800 Ma is much like that cited above, except that behavior joins morphology, life style, and environment in interpretation. Eohyella, in fact, occurs in 1700-Ma stromatolites from China (Zhang and Golubic 1987), but specific comparisons to modern species cannot be made at present." Knoll and Golubic (1992:456,457).
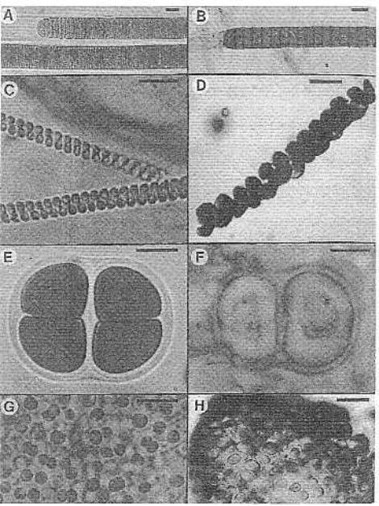
Living stromatolite-building cyanobacteria from northern Mexico (A, C. E, and G) and Precambrian look-alikes (B, from the 950-Ma-old Lakhanda Formation, and D, the 850-Ma-old Miroedikha Formation, both of Siberia; F, the 1,550-Ma-old Satka Formation of Bashkiria; and G, the 2,100-Ma-old Belcher Supergroup of Canada, Hudson Bay. (A) Lyngbya, compared with (B) Palaeolyngbya. (C) Spirulina, compared with (D) Heliconema. (E) Gloeocapsa, compared with (F) Gloeodiniopsis. (G) Entophysalis, compared with (H) Eoentophysalis. Bars for scale represent 10µm. From J. W. Schopf (1999) Fig. 8.10.
During these billions (thousands of millions) of years, these one-celled organisms have not evolved at all into any higher forms of life. They do not know anything about evolution. Since they began, they have been perfectly adapted to their environment, to their specific niche in life. The geological record, the sediments laid down during the last 4 billion (4000 million) years, do not know anything about evolution.
How complex were the old one-celled organisms, whose fossil remains one has found now in different parts of the world? How much information have the bacteria and archaea contained, which have lived long before us on our planet earth, billions of years ago? How long was their genome (DNA-chain)? And much information was needed, to put its nucleotides into the right order? In other words: What are the sequence alternatives for their genome, so that each base pair was put at the right place?
Beggiatoa. At Warrawoona, West Australia, the fossil remains of the bacterium Beggiatoa arachnoidea and Beggiatoa minima have been found. They have lived there some 3.3-3.5 billion years ago (Schopf, J. W. 1983, Table 9.2). Most of the cell's genetic information is encoded in its genome, on its DNA-chain. How long was their genome? And how much information was needed, to put its nucleotide pairs into the right order?
We can find this out only on the genus level, since so far only the genome size of Beggiatoa alba has been published. It has 2.02·109 Da = 3.03·103 kilobase pairs, according to Genthner et al., 1985, Bergey's Manual (1989:2092).
2.02·109 Da : 660 Da/bp = 3 060 606 bp
3 060 606 log 4 = 101 842 668 bit information, sequence alternatives.
This means: At least 101 842 668 bit information (yes/no decisions) were needed, to put the base pairs in the DNA-chain of the genus Beggiatoa into the right order. We should remember here: All of mankind's knowledge, written down now into books, is "only" 1018 bit (Gitt, W. 1986:68). The differences within a genus of a bacterial cell are only racial variations of their basic kind. That has nothing to do with the evolving into higher kinds of plants and animals.
Methanococcus. At Warrawoona, Western Australia, one has also found the fossil remains of the archae Methanococcus vaniellii. It has lived there some 3.3-3.5 billion years ago (Schopf, J. W. 1983, Table 9.2). The genome size of two other species of Methanococcus has been published, of M. thermolithoautotrophicum and M. voltae. Methanococcus thermolithoautotrophicum SN 1 has a genome size of 1.1·109 Da, as reported by A. Klein and M. Schnorr (1984:630).
1.1·109 Da : 660 Da/bp = 1 666 666 bp log 4 = 101 132 044 bit.
The genome of Methanococcus voltae has 1 870 000 to 1 899 000 base pairs, and a mean of 1 880 286 bp, as reported by Sitzman, J. and A. Klein (1991:505-513).
1 880 286 bp log 4 = 101 132 044 bit.
This means: If we take the lower value of 101 003 432 bit, it took at least 101 003 432 bit of information (or yes/no decisions) to make the DNA-chain of the archae Methanococcus. So much information was needed, just to put the base pairs of its genome into the right order, some 3.5-3.5 billion years ago! From where has this information come?
Cyanobacterium. The bacteria have not evolved at all into higher forms of life. They have been perfectly adapted to their environment since the beginning. Bacteria, whose fossilized remains one has found in 3.3-3.5 billion year old sediments, can even be identified according to their genus, and some even according to their species. That is of genera and species, which are still living today. If they had evolved through mutation and selection, as evolutionists still want us to believe, what should we expect then? Should we not expect then, that for each suitable bacterial cell, we should find hundreds or thousands of unsuitable cells, in all their different stages of evolution?
Should we not expect then that the sediments of our earth contained mostly bacterial scrap, like a huge garbage dump? But we only find instead perfect cells since the beginning, no biological garbage, nor any intermediate stages. Nor do we find any trace of a primitive "common ancestor of all life". It exists only in the fantasy of deluded evolutionists. They have no experimental nor observational proof whatsoever for their assertion. It is only a pious myth.
Not only the oldest fossilized cells, some 3.5 billion years old, which we know now, but also these found later on in younger sediments, 2 billion and 1 billion years old, are just as perfect. Many of them can be identified with species, which are still living today. Let us look now briefly at a few cyanobacteria.
Lynbya. Prof. J. W. Schopf compares the detected Warrawoona cells with those, which are still living today. Among the many different genera and species of bacteria (and archaea), he mentions also the oxygen producing cyanobacterium Lynbya cryptovaginata. It is 3.3-3.5 billion years old. It has a diameter of 4.0-9.0 µm. At Warrawoona, also five different species of the cyanobacterium Phormidium have been found. They have a diameter of 0.6-6.7 µm, and a length of 2.5-8.0 µm. Schopf, J. W. (1983) table 9.2. They are grouped now with Lyngbya. Mann, N. H. et al. (1992:34).
In the Lakhanda Formation of the Khabarovsk region of eastern Siberia, Palaeolyngbya, a Lyngbya-like fossil cyanobacterium, about 950 million years old, has been found. Compare, please, figure 2.8A with 2.8B in Schopf, J.W. (1992:51). How large is the genome of the oxygen producing cyanobacterium Lyngbya? How much information does it contain?
The genome size of Lyngbya (strain PCC 7419) is about 4.58·109 daltons, as reported in Bergey's Manual (1989:1777), and Herdman, M. et al. (1979b). 4.58·109 Da : 660 Da/bp = 6 939 393 bp log 4 = 104 177 930 bit.
Spirulina. This is an oxygen producing cyanobacterium. Spirulina remains, 850 million years old, one has found in the Miroedikha Formation of the Turuchansk region of eastern Siberia. See, please, pictures in Figure 2.8C and 2.8D in J. W. Schopf (1992:51). How large is its genome? Spirulina has a genome size of 2.53·109 to 5.19·109 Da, and an average 3.86·109 Da. 3.86·109 Da : 660 Da/bp = 5 848 484 bp log 4 = 103 521 138 bit.
Genome size from M. Herdman et al. (1979:80). Thus, it takes 103 521 128 yes/no decisions to make the DNA chain of this cell. That is, to put its base pairs into the right order. Chance, mutation, and selection cannot do that.
Gloeocapsa. Gloeodiniopsis is a Gloeocapsa-like fossil cyanobacterium about 1 550 million years old from the Satka Formation of the southern Ural Mountains of Bashkiria. See, please, pictures of old and new cell in Schopf, J. W. (1992:51). Figure 2.8E and 2.8F. It has not evolved at all. The fossils clearly disprove the doctrine of evolution. They expose it as a pious myth. How large is the genome of Gloeocapsa? How much information does it contain?
The genome size of Gloeocapsa is 2.90·109 to 3.47·109 Da. Its average is 3.20·109 Da, as reported by M. Herdman et al. (1979:77). 3.20·109 Da : 660 Da/bp = 4 848 484 bp log 4 = 102 919 078 bit.
This means: 102 919 078 yes/no decisions were needed, to put the base pair of its genome into the right order. This is like putting the letters and words of a written text into the right order. How much information is that? We shall understand this a little better, if we remember, how much information mankind has written down till now into books. "Only" 1018 bit (W. Gitt, 1986). Chance cannot make any information. But it can destroy information. The more mistakes there are in a computer program, the less useful it will become. Also thousand apes, writing on their typewriters one of Shakespeare's poems, do not produce any information at all. Important is here only the producer (and sender) of the information, not the receiver (M. Himmelheber).
From the time, when our planet Earth arose, some 4.55 billion years ago, to the time, when one finds the first one-celled organisms, about 3.8 billion years ago, about 700 000 million years went by. During these 700 000 million years, the first living cell as the "common ancestor of all life on Earth" is supposed to have evolved by itself from inorganic matter. But for this belief, there is no sedimentary proof at all. During these 4 billion years, since one-celled organisms live on this Earth, they have not evolved at all into any higher forms of life. The first living cell was just as complex, as those living today. The cell itself has not "discovered" or "invented" anything. The chemical systems, which they have used then, they are still using today. Some of these first cells can be identified according to the genus, or even to the species of one-celled organisms, still living today. They have not evolved at all into any higher forms of life. That is just a pious, religious myth. It has nothing to do at all with serious science.