
Chapter 4: New Precambrian Findings
What have scientists found out now about the oldest one-celled organisms at the beginning of the Precambrian Time? How have they come into being? How complex were they? Have they really come from a common ancestor, the "first ancestral cell"? And have these bacteria then evolved into higher forms of life, into plants, animals, and human beings? Is the evolutionary "Tree of Life" fact or fiction?
Preston Cloud is Professor emeritus of Geology at the University of California at Santa Barbara. He is a member of the National Academy of Sciences. And he was associated with the United States Geological Survey. He believes in evolution - Why is there life on Earth? How has it arisen? How complex was it?
Prof. Preston Cloud: "Conclusions about early surface conditions that emerge from these observations (in Archaean time) are: By no later than 3.8 aeons ago, a solid, emergent crust was already present and on it a variety of sediments and interlayered volcanic deposits. An atmosphere rich in carbon dioxide and water vapor was also present at that time, moderating global temperature and accounting for the weathering manifested by the existence of sedimentary rocks.
"An upper limit of 86°C to 146°C has been derived from oxygen-isotope ratios of the chert, and computer modeling suggests a surface temperature of 85° to 110°C. As the chert (at Isua, SW Greenland), however, is probably an altered volcanic ash, atmospheric temperatures were most likely not above boiling. The evidence for flowing surface waters given by the Isuan gravel deposits makes it unlikely that prevailing surface temperatures differed greatly from present ones, although it was perhaps uncomfortably warm by human standards." (1989:133, 134).
How complex were the first organisms, that arose on the Earth, some 4 billion years ago, when its surface had cooled down enough?
Prof. Preston Cloud: "The simplest organisms were already more complex than the most complicated physical systems we know. ... The oldest possible fossil remains, however, have the sizes and shapes of bacteria, and the oldest indirect evidence for life suggests microbial agents. ... If living today, they would surely be identified as bacteria (or perhaps archaebacteria). A shallow, anoxic marine habitat, shielded from radiation by water or mud, or both, would have been a likely starting site.
"Compared with known living organisms it could hardly have been called anything but a bacterium. Such wee creatures may not look very clever, but they are extraordinarily inventive biochemically. They have explored every conceivable habitat, and some inconceivable ones - like sulfuric acid and carbolic acid, smouldering coal, and ... jets of hydrogen sulfide-rich fluids at the crests of deep-oceanic spreading ridges. They outwit mankind's most elaborate efforts to exclude them from their company. And one may be sure that their ancestors were a match for the monotonously anoxic habitats of Archaean history. Bacteria even evolved the ability to repair molecular dislocations in their DNA caused by high-energy ultraviolet radiation - persuasive evidence for their descent from lineages that existed before there was enough O2 to create a shielding ozone screen." Cloud, P. (1989:42, 233, 234).
Manfred Schidlowski is a Professor at the Max-Planck Institute for Chemistry, in Mainz, Germany. He is one of the foremost experts on Precambrian fossils in the world. He believes in evolution. What has he found out about the first living cell on earth, and how it has come into being? He says in Instruments, Methods, and Missions for the Investigation of Extraterrestrial Microorganisms, 29 July-1 August 1997, San Diego, California, under the heading, "Carbon isotopes and the oldest record of life: Potential and limits":
"The currently available sedimentary carbon isotope record goes back to 3.85 Ga and conveys a remarkably consistent isotopic signal of biological carbon fixation based on the bias for light carbon (12C) exercised by common photosynthetic pathways. This holds particularly for the time segment < 3.5 Ga (=3.5 billion years). ... A cogent case can be built for the emergence of (photo)autotrophic carbon fixation and the start of a biogeochemical carbon cycle as from at least 3.85 Ga ago. This would imply that microbial (procaryotic) ecosystems had been prolific on the Archaean Earth not long after the formation of the planet." (1997:462).
"It is by now firmly established that there exists a continuous record of organic carbon (Ccarb) in sedimentary rocks that can be traced back to the 3.8 Ga-old Isua Supracrustal Suite in West Greenland. ... A straightforward implication of these findings is that biological carbon fixation (and notably photoautotrophy) attested by the Archaean stromatolite record, prokaryotic mirobenthos demonstrably held dominion over the Earth as from at least 3.5 Ga ago and possibly earlier. It should be noted also that microbial communities figure among the most productive ecosystems of the extant biosphere, with specifically benthic prokaryotes (such as cyanobacteria) capable of sustaining prodigious rates of primary productivity on the order of 10 g Corg/m² day. If such high rates of primary production can be maintained by microbial photoautotrophs operating on the prokaryotic level, photosynthesis may have gained little in quantitative importance during subsequent evolution of life." (1977:466, 467)
"Over 3.5 Ga of the unmetamorphosed rock record, isotope fractionations between carbon and carbonate thus appear to be demonstrably the same as in the present world... There is little doubt that the glaring 12C-enrichment displayed by the data envelope for fossil organic carbon constitutes a coherent signal of autotrophic carbon fixation over 3.85 Ga of Earth history as it ultimately rests with the process that gave rise to the biogenic precursor materials. Moreover, the basic uniformity of this isotopic signal through time attests to an extreme degree of conservatism in the principal biochemical pathway of carbon fixation. In fact, the mainstream of the envelope for d13Corg depicted in Fig. 3 (in his article) can be most readily explained as the geochemical manifestation of the isotope-discriminating properties of one single enzyme, namely, ribulose-1,5 bisphosphate (RubP) carboxylase, the key enzyme of the Calvin cycle (found in Cyanobacteria, Purple bacteria, and in plants). Hence, the d13Corg age function may be aptly termed an index line of autotrophic carbon fixation over almost 4 billion years of recorded Earth history, suggesting that prokaryotic ecosystems had been established on the Archaean Earth not long after the formation of the planet." Schildlowski, M. (1997:468).
In "Instruments, Methods, and Missions for Astrobiology", 20-22 July 1998, San Diego, Professor Manfred Schidlowski states under the heading "Beginnings of terrestrial life: Problems of the early record and implications for extraterrestrial scenarios":
"With the paleobiological evidence from the currently known Archaean rock record at hand, the existence on Earth of microbial (prokaryotic) ecosystems as from about 3.8 Ga ago seems so firmly established as to be virtually unassailable. ... (There are) problems of the time scale of early organic evolution as exemplified by the unheralded appearance about 3.8 Ga ago of microbial life on the organizational level of the prokaryotic cell." (1998:149).
"With empirical evidence regarding the initiation of life processes of the early Earth virtually cut off along with the truncation of the sedimentary record some 3.8 Ga ago, the very beginnings of life on this planet keep being shrouded in mystery." (1998:149)
"Considering the remarkable degree of morphological and possibly also physiological diversification that characterizes the oldest well-preserved microbial communities (notably the Warrawoona microflora of Australia with an age just short of 3.5 Ga) it should go without saying that the ancestral lines of bacterial (prokaryotic) life must extend well back into the time segment covered by the metamorphosed record. With the existence of precursor floras to the Warrawoona community thus almost certain, reports of cell-like morphologies from the isual metasediments were bound to attract considerable attention. This was particularly true for the key morphotype of the Isua assemblage, Isuasphaera isua, a globular and supposedly sheath-enclosed cell-like microstructure recovered from a metachert horizon." (1998:151).
About "The Problem of the missing Archaebacterial Record", Prof. M. Schidlowski then says: "Since the majority of cyanobacterial taxa can be clearly identified and distinguished from other members of the microbial domain by their morphological characteristics (form, cell size and attendant organizational/structural detail), the micropaleontological morphotype inventories of the oldest rocks suggest that cyanobacterial ecosystems have held dominion over the early Earth. In fact, the Archaean-Proterozoic record is dominated by evidence on the cyanobacterial level, while members of the archaebacterial lineage are obviously missing. This is the more surprising since Archaea in the widest sense show up prominently at the very base of terrestrial life.
"Molecular phylogeny (= evolution) based on the assessment of nucleic acid sequences permits the inference that all terrestial life is ultimately derived from a single common ancestor or 'progenote', with subsequent diversification proceeding in an evolutionary continuum that is grounded on Darwinian evolution and operates on a genetic program relying on DNA. In contrast to such postulates, the earliest paleontological record is characterized by evidence on the cyanobacterial level, even in lithologies formed in close proximity to high-temperature environments such as the microfossil-bearing cherts within the ~3.5 Ga-old Apex Basalts of the Warrawoona Group of Western Australia." Schidlowski, M. (1998:153)
"Even a dense pounding by small- to medium-scale asteroids of the Earth as a whole could not have led to a complete annihilation of early terrestrial life as long as vaccination strains were able to survive in protected niches or unaffected terrains. Accordingly, there is good reason to believe that specifically the late-stage asteroid bombardment of the juvenile planet was unlikely to ever have seriously interfered with the early evolution of prokaryotic life."
What about the "Problems with the Time Scale of Early Organic Evolution"?
Prof. M. Schidlowski: "With the current available evidence at hand, a good case can be built that microbial (prokaryotic) life was firmly entrenched on the ancient Earth as from at least 3.8 Ga ago (Fig. 1), with the concomitant emergence of a biologically modulated bipartite carbon cycle comprising a reduced (organic) carbon) and an oxidized (carbonate) branch. Considering the age of 4.55 Ga for the planet as a whole, any Earth-based origin of life and its subsequent evolution to the prokaryotic level must have been accomplished within the uncomfortably short time of 0.7 Ga." (1998: 154,155).
Comment: In this time, the surface of the Earth was still so hot that there has been at first no liquid water at all. And later, when it had cooled off a little, it was covered with hot, boiling water. Hence, at this time, no life was able to evolve at all by itself from inorganic matter.
Photosynthetic bacteria, using the Calvin cycle, have lived on this earth since the beginning of the sedimentary record. They have produced so much free oxygen in a reducing environment, that human beings and animals are also able to live now on this planet. How complex was this Calvin cycle, with its key enzymes? What did these photosynthetic bacteria, 3.6-3.8 billion years ago, all have to know about physics and chemistry? What have scientists found out about this now? The bacteria had to know more about microbiology, physics, and chemistry, than any scientist, now living on this planet earth. Also the oldest one-celled organism is a fully automated chemical factory. It is so complicated, that no human being is able to make it. If you, dear reader, do not understand some or most of the following technical terms, do not worry about this. It will show you then still clearer, how much the first living cell on earth had to know about science, and what it was able to do.
Michal T. Madigan, John M. Martinko, and Jack Parker are professors of microbiology at the Southern Illinois University-Carbondale, USA. They believe in evolution. They state in their textbook, Biology of Microorganisms (1997:488) about Key enzymes of the Calvin cycle:
"The first step in CO2 and ribulose bisphosphate leading to the formation of two molecules of 3-phosphoglyceric acid (PGA), one of which contains the carbon atom from CO2. PGA constitutes the first identifiable intermediate in the CO2 reductive process. The carbon atom in PGA derived from CO2 is still at the same oxidation level as it was in CO2, and the next two steps involve reduction of PGA to the oxidation level of carbohydrate. In these steps, both ATP and NADPH are required: the former is involved in the phosphorylation reaction that activates the carboxyl group, the latter in the reduction itself.
"The average carbon atom in glyceraldehyde phosphate is now at the reduction level of carbohydrate (CH2O), but only one of the carbon atoms of glyceraldehyde phosphate has been derived from CO2, the other two having arisen from the ribulose bisphosphate. However, this is only the first part of the Calvin cycle. Most of the remaining reactions involve rearrangements to regenerate ribulose bisphosphate molecules, with some of the recently fixed CO2 going to new cell synthesis."
"The series of reactions leading to the synthesis of ribulose bisphosphate involve a number of sugar rearrangements. Through the action of enzymes that rearrange pentose phosphate compounds and enzymes of the glycolytic pathways, glyceraldehyde 3-phosphate is converted to ribulose 5-phosphate and subsequently to ribulose bisphosphate.
"It is best to consider reactions of the Calvin cycle based on the incorporation of 6 molecules of CO2. For RubisCO to incorporate 6 molecules of CO2, 6 ribulose bisphosphate molecules are required as acceptor molecules. This yields 12 molecules of 3-phosphoglyceric acid (a total of 36 carbon atoms). These 12 molecules serve as carbon skeletons to form 6 new molecules of ribulose bisphosphate (a total of 30 carbon atoms), and 1 molecule of hexose for cell biosynthesis. A complex series of rearrangements involving C3, C4, C5, C6 and C7 intermediates finally yields 6 molecules of ribulose 5-phosphate, from which the 6 ribulose bisphosphates are generated. The final step in the regeneration of ribulose bisphosphate is the phosphorylation of ribulose 5-phosphate with ATP by the enzyme phosphoribulokinase. This enzyme is another catalyst unique to the Calvin cycle.
"Let us consider now the overall stoichiometry for conversion of 6 molecules of CO2 into 1 molecule of fructose 6-phosphate. Twelve molecules each of ATP and NADPH are required for the reduction of 12 molecules of phosphoglyceric acid (PGA) to glyceraldehyde phosphate, and 6 ATP molecules are required for conversion of ribulose phosphate to ribulose bisphosphate. Thus, 12 NADPH and 18 ATP are required to synthesize 1 hexose molecule from CO2. Hexose molecules can be converted to storage polymers such as glycogen, starch, or poly-ß-hydroxyalkanoates during periods when ATP and NADPH are abundant and then can be used in other periods, such as in darkness, as sources of carbon and energy.
"The key enzymes of the Calvin cycle, ribulose bisphosphate carboxylase and phosphoribulokinase, are unique to autotrophs that fix CO2 via the Calvin cycle. These enzymes have been found in virtually all phototrophic organisms examined - plants, algae, and bacteria. They are also found in many chemolithotrophic Bacteria, such as the sulfur, iron, and nitrifying bacteria." Madigan, Martinko, Parker (1997:488-489).
He is a member of the Department of Earth and Space Sciences, the Molecular Biology Institute, and the Institute of Geophysics and Planetary Physics (IGPP) at the University of California, Los Angeles. He is Professor of Paleobiology and Director of the IGPP Center for the Study of Evolution and the Origin of Life. And he is a member of the National Academy of Sciences of the USA. He believes in evolution. In the last chapter, we studied his earlier reports about the first living cells. We would like to find out now: In what kind of an environment have the first cells lived? How have they come into being? Since when have there been cyanobacteria on this earth? How were these photosynthesizing bacteria able to live, when there was yet no free oxygen and no ozone shield? He reports in his new book, Cradle of Life (1999:88, 89), about the Apex chert and its fossils in northwestern Australia:
"The Apex fossils are preserved in a chert bed sandwiched between two massive lavas of the Pilbara sequence. Zircons in a lava immediately overlying the fossil bearing chert have a U-Pb age 3,485 ± 1.9 Ma, whereas those in a lava below the chert are 3,471 ± 5 Ma in age. The fossil-bearing rock unit is therefore older than about 3,460 Ma and younger than about 3,47 Ma; the bed has an age of 3,465 ± 5 Ma. ... The rock unit that contains the Apex fossils has an age of 3,465 ± 5 Ma, the fossils themselves are actually even older. The fossils are preserved in small rounded granules, a millimeter or so in size, that are embedded in a type of rock known as a grainstone conglomerate and made up of many such rocky chunks.
"Conglomerates like the Apex chert are formed along wave-washed beaches and at the mouths of streams and rivers. The granules and pebbles that make up the deposit are bits and pieces of rocks originally formed someplace else and then broken up and carried by flowing water to the Apex bed. Only one of the many types of rounded stones in the conglomerate contains fossils. Their well-rounded sides and small size shows that they were transported a long distance, but no one knows from where. Unless the 'mother lode,' the source bed, can be found and dated, we will never know the full age of the Apex fossils. They are older than 3,465 ± 5 Ma, but how much older remains a question."
"The scene was dominated by broad, shallow seaways, into which volcanic lavas erupted. Scattered volcanic islands were fringed by river gravels, sandy inlets, mudflats, and occasional evaporitic lagoons. The Apex chert occurs within one such shallow-water-sequence, sandwhiched between two massive lava flows, on the western flank of what is known as the Mount Edgar Batholith. The Australian Pilbara and South African Swaziland rocks, the only thick geologic sequences known to have survived from this distant time, are made up mostly of greenstone belts. ... But as the only remnants we have to go on, they paint a picture of wide shallow seas dotted with abundant rocky volcanic islands and their smoldering fumeroles and hot springs." (1999:90, 91).
What kinds of organisms have lived there?
Prof. J. W. Schopf: "All of the Apex fossils are remnants of cellular filamentous microbes known as prokaryotes, an early evolving type of microorganism in which the hereditary material (DNA) is in simple strands within the cell rather than packaged in a cell nucleus as in more advanced forms of life (eukaryotes). Among the prokaryotes, all of the Apex fossils belong to the domain known as Bacteria (which includes cyanobacteria as well as less advanced bacterial types) rather than the Archaea, the other prokaryotic domain (a rather recently discovered branch of the Tree of Life made up of microbes that often live in harsh, high-temperature, acidic settings).
"In fact, several of the larger Apex species so closely resemble modern and Proterozoic cyanobacteria that it seems to me likely they are members of a common and particularly well-known subgroup that still lives today (a formal taxonomic family of cyanobacteria, the Oscillatoriacea).
"If I am right about these relations, the presence of cyanobacteria in this nearly 3,500-Ma-old community tells us that early evolution proceeded very far very fast. All cyanobacteria are able to do the kind of photosynthesis that gives off oxygen; and, like higher plants and animals, all can breath in oxygen (by a process known as aerobic respiration). Both of these, however, are advanced ways to live, evolved from more primitive ways in which free oxygen plays no part.
"So, if cyanobacteria existed at this early time, the earlier evolved processes must also have been present; the living world would have to have included organisms that photosynthesized without giving off oxygen (bacterial photosynthesizers) as well as those that produced it (cyanobacteria), and microbes that lived in the absence of oxygen (anaerobes) as well as those that breathed it (aerobes). These are precisely the same processes that power the present-day living world. If cyanobacteria are represented among the Apex fossils, we are forced to conclude that the basics of the world's ecosystem had already become established by this early stage in the history of the Earth.
"No one has publicly disagreed with my interpretation of the Apex fossils. But, privately, some would prefer I were mistaken, since they (and I, too) would prefer a simpler evolutionary story, one that told us these oldest fossils organisms were capable only of primitive ways of living and that advanced metabolic lifestyles evolved much later. But the evidence seems strong, and what one might 'prefer' shouldn't matter."
"Many prokaryotes and almost all cyanobacteria secrete mucilage from their cells, but the Apex community is the only one known where the microorganisms lived embedded in such massive clumps. ... But it could be that this sticky secreted mass glued the Apex organisms to the shallow seafloor, where it enabled them to harvest sunlight protected from damaging UV rays by overlying seawater. If so, like other traits characteristic especially of cyanobacteria (their ability to glide away from intensive light and to manufacture biochemicals both to protect themselves from harmful UV and to repair the damage it causes), the extracellular glue played a role in helping these early evolving microorganisms cope with an inhospitably harsh environment."
"The oldest fossils ... are eleven kinds of cellular threadlike microbes petrified in the 3,465-Ma-old Apex chert of northwestern Western Australia. These are recent finds on which further study is needed. But it already seems clear that all are prokaryotes of the Bacterial domain and the best evidence is that the fossils include several types of oxygen-producing and oxygen-breathing cyanobacteria. If so, these organisms are not only extremely ancient but surprisingly advanced, and show that early evolution proceeded faster and farther than anyone imagined.
"Even the simple act of breathing - taking in oxygen and using it to burn foodstuffs to generate energy - involves many steps, each controlled by a different enzyme. As in breathing, each event in every biochemical process is governed by a different enzyme. Life requires so many enzymes that most of the chromosomal genes are earmarked for their production. ... Breathing oxygen via aerobic respiration is a vast improvement over earlier-evolved glucose fermentation (glycolis alone). The primitive process makes two ATPs from each glucose metabolized - the equivalent of only 2% of the energy stored in each molecule - whereas the oxygen-using system yields thirty-six, a whopping 38% of the energy available (and a yield better than the 25% efficiency of most automobile engines."(1999:96-100, 104, 158, 159).
Can the evolutionary "Tree of Life" tell us, when the first cells have arisen on earth?
Prof. J. W. Schopf: "Accurate dating of life's early branching remains a challenge. There are good reasons to hope that the molecular makeup of living cells hold the key, but neither rRNA trees nor dates based on evolving proteins has yet proved reliable. Likewise, early fossils are still too incompletely known to provide precise answers and, in any event, can record only the first detected occurrence of a biological group, not its actual presence." (1999:106, 107).
There are two kinds of living organisms: Those that live by making their own food from inorganic matter. And those that live by eating other organisms. Which type of living is more complicated?
Prof. J. W. Schopf concludes: Organisms (like animals) are metabolically simpler than plants. Those who eat others, use ready-made foodstuff and only need to break it down. But plants and other autotrophs must make their own food and break it down as well. (1999:145). - Thus, the first cell, living on inorganic food, had to be more complex, than the one, who arose later on, and who ate the remains of other bacteria. That is right the opposite of what we would expect according to the hypothesis of evolution.
Flattened filaments in siltstones. Compressed Precambrian filaments are especially well known from shaley rocks of Siberia (A and B, from the 850-Ma-old Miroedikha Formation; C, the 950-Ma-old Derevnaya Formation) and Bashkiria (D, from the 925-Ma-old Sim Formation; E and F, the 925-Ma-old Shtandin Formation). (A) Primorivularia; (B) Trachythrichoides; (C) Partitiofilum; (DI Heliconema; (E and F) Calyptothrix. Bars for scale represent 10 µm. From J. W. Schopf (1999) Plate 3.
What are now the oldest fossils in the world? When have they arisen?
Prof. J. W. Schopf: "The oldest fossils known - tiny cellular threads entombed in the Apex chert of northwestern Australia - are nearly three-quarters the age of the Earth. The fossil-bearing bed was laid down along the edge of a narrow seaway flanked by soaring volcanoes that episodically blanketed the seafloor with massive lava flows. Sandwiched between two of these, each precisely dated, the fossiliferous horizon is 3,465 ± 5 Ma old.
"Several types resemble living cyanobacteria of the taxonomic family Oscillatoriacea, today an especially common group shown by rRNA evolutionary trees to be one of the most primitive kinds. Their relation to this particular family of cyanobacteria also meshes with the younger, Proterozoic, Precambrian fossil record, where oscillatoriaceans preserved in the same way in similar settings and microbial communities are widespread and abundant. ... It fits well with the chemistries of ancient minerals and organic carbon, which show that cyanobacterial photosynthesis may actually date from even earlier than 3.5 billion years ago.
"Cemented to rocks and boulders on the seaway floor by a thick layer of sticky mucilage and protected by overlying waters from lethal UV light, the Apex community was evidently composed of cyanobacteria as well as other kinds of prokaryotes, and included autotrophs (= self-feeders) and heterotrophs (= living on other bacteria) and both anaerobes and aerobes (the last, probably 'facultative' like some oscillatoriaceans and many other microbes that consume oxygen when available but otherwise live by anaerobic means).
"The origin of life cannot be dated precisely. The Apex fossils set a minimum age, yet are too varied and advanced to be close to life's beginnings. Living systems arose earlier than 3,500 Ma ago, during the first billion years of Earth's existence, but just when is an open question. ... But life as we know it could come into being only after 3,900 Ma ago, and by a scant 400 Ma later it was flourishing and widespread. How did life advance so far so fast?" (1999:164-167).
What geological evidence is there that there were cyanobacteria producing free oxygen?
Prof. J. W. Schopf: "Of all microbes, only cyanobacteria carry out oxygen-producing photosynthesis, a process that of course cannot take place without its two starting ingredients, water and carbon dioxide, that forms two products, organic matter and molecular oxygen. Evidence of all four should be unmistakably in the rock record once cyanobacteria appeared.
"There is ample proof of liquid water. The fossil-bearing Apex chert is part of a sequence of volcanic and sedimentary rock some 15 kilometers thick, nearly all deposited in shallow seas. Its geology tells us the scene was dominated by sinuous seaways with marginal lagoons and scattered volcanic islands fringed by water-laid gravels, sands, and muds. The other ingredient needed for photosynthesis, CO2, was also plentiful. Carbon dioxide solubilized in seawater as bicarbonate (HCO3-) combined with dissolved calcium (Ca+2) to form the calcium carbonate (CaCO3) minerals that make up limestone beds of the Apex sequence..
"The two end products of cyanobacterial photosynthesis, organic matter and O2, were present as well. Particles of coaly carbonaceous matter, kerogen, are abundant in the Apex rocks (constituting up to about 1% of their weight) and make up the carbon-rich cell walls of the petrified fossil microbes. The presence of free oxygen is shown by iron-oxide-rich banded iron formations (BIFs)." (1999:170, 171).
How have the banded iron formations arisen? Why are they in layers? And why are they red? Why have they arisen?
Prof. J. W. Schopf: "BIFs are the principal source of the world's iron ore. Their distinctive millimeter- to centimeter-thick banding is caused by alternation of iron-rich and iron-poor layers; because the iron is in the form of tiny rustlike rains of hematite (Fe2O3) and sometimes magnetite (Fe3O4) they have a telltale bright to dull red color.
"The iron owes its origin to the circulation of seawater through hot cracks and fissures in the ocean floor, primarily at the deep submarine ridge systems that play an active role in plate tectonics. In a dissolved form (as ferrous, divalent iron, F+2) it then spread upward into shallower reaches of the water column - often seasonally, giving rise to the distinctive fine banding. There it was oxidized to the ferric state (trivalent iron, Fe+3), chiefly by combination with dissolved molecular oxygen, and rained out of solution as a rusty mist of minute iron oxide particles.
"But an enormous amount of oxygen - more than 20, 000 million trillion grams (2·1022 gm), roughly twenty times as much as in the atmosphere today - is buried in the iron oxides of BIFs. Of all sources, including nonbiologic ones (such as the breakdown of water vapor by intense heat or high energy UV light), only photosynthesis is capable of generating such a massive amount.
"BIFs are abundant over a long span of geologic time, from earlier than 3,500 Ma to about 2,000 Ma ago, and are mined for steel worldwide, in Australia, Africa, Asia, Europe, North and South America. They are a testament of a truly striking stage in planetary history - the rusting of the Earth - which drew to a close only when the ocean was finally swept free of dissolved iron. Yet the presence of these remarkable deposits does not mean the oceans were oxygen rich. On the contrary. BIFs were nearly always deposited in large basins, hundreds of kilometers in length and breadth, and the dissolved ferrous iron from which BIFs form could be spread over such expanses only if carried by waters that were oxygen poor.
"Huge amounts of molecular oxygen were pumped into the environment by oxygenic (cyanobacterial) photosynthesis, but except locally, near where it was produced, amounts of oxygen were kept low by its capture and rapid burial in the oxide minerals of BIFs. ... Evidence both from fossils and from geology points to the existence of oxygen-producing photosynthesizers, cyanobacteria, by 3,500 Ma ago. ... Like the evidence from cellularly preserved fossils, carbon isotopes show that photosynthetic members of the Bacterial domain, probably cyanobacteria and photosynthetic bacteria, both, have existed since 3,500 Ma ago. And measurements on graphitic carbon from the 3.8 billion-year-old Isua sequence hint that photosynthesizers may have been present then too." (1999:170-173, 177).
Stromatolites are layers, in which bacteria have lived. In the highest layer, at its surface, the photosynthesizing cyanobacterium has lived. Below it, other kinds of bacteria. Since when are there stromatolites? Have they evolved into higher forms? And have the bacteria, which built them, evolved during the billions of years into higher forms of life?
Prof. J. W. Schopf: "Scores of specimens are in places packed together cheek by jowl in a horizon of the Western Australia Pilbara sequence (dated to be 3,450 Ma old and therefore arguably the oldest stromatolitic bed known) that extends over a reported 'tens of square kilometers,' a spread much too broad to be accounted for by localized soft-sediment deformation. ... The reefs of domical and columnar ones at Shark Bay closely resemble fossilized reefs in South Africa 2,300 Ma old, half the age of the planet; specimens of one-billion, two-billion, and more than three-billion-year-old columnar stromatolites all look nearly the same; and the conical kinds, less common than others, seem not to have changed much, either. What this means is that stromatolites can tell us about past environments, but probably not very much about evolution.
"Even though stromatolites didn't evolve, one would think the micro-organisms that built them, must have giving rise to what can be called a 'Volkswagen Syndrome' - a lack of change of external form that masks internal evolution of the working parts. This can be checked by examining the microscopic fossils petrified in ancient stromatolites, the cellular and tubular filaments and single-celled and colonial spheroids that make up their mats, and comparing these to microbes that build mats today. ... We have already seen a couple of well-preserved microbial mat-building communities - those of 850-Ma-old flat-layered stromatolites of the central Australian Bitter Springs Formation and 2,100 Ma-old domical stromatolites of the southern Canadian Gunflint chert.
"We were introduced to a third assemblage, likely stromatolitic too ... from the nearly 3,500-Ma-old Apex chert of northwestern Australia. But these three benchmark communities, known for their contributions to the development of the science and understanding of life's early history, are only a tiny sample of more than 250 that have been found in Precambrian stromatolites worldwide, a list of locales that reads like the United Nations - Australia, Brazil, Canada, China, France, India, Israel, Kazakhstan, Norway (Svalbard), Russia, South Africa, the USA.
"So, there are now plenty of examples of fossil communities that one can compare - like the stromatolites they build, the mat builders themselves evidently didn't evolve, either! This absence of change seems so contrary to the usual view of Darwinian evolution that it needs to be fully documented and explained before it can be excepted. Suffice it here to say that there is strong evidence that the same kinds of organisms living in the same kinds of environments built the same kinds of stromatolites over billions of years." (1999:197, 201).
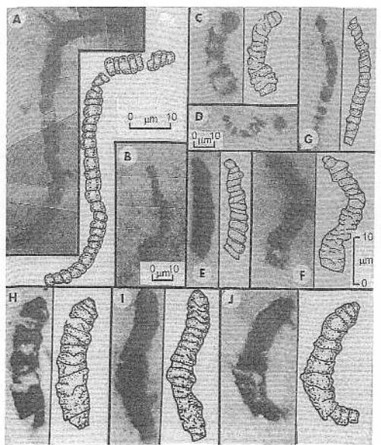
The oldest fossils, cellular cyanobacteriumlike filaments shown in thin sections of the 3,465 ± 5-Ma-old Apex chert of northwestern Australia. Because the petrified microbes are three dimensional and sinuous, composite photos have been used to show the specimens in (A), (B), (D), and (F through J). (A and B) Primaevifilum amoenum, a species having cylinder-shaped body cells and rounded end cells; (C through F) Archaeoschillatoriopsis disciformis, tapered filaments composed of disk-shaped body cells; (G) Primaevifilum delicatulum, a narrow species of the genus Primaevifilum; (H, I, and J) Primaevifilum conicoterminatum, a species that has distinctive conical end cells. The 10-µm-long scale in (F) shows the size of all fossils except as otherwise indicated in (A), (B), and (D). From J. W. Schopf (1999:77) Fig. 3.4.
Since when has one known Precambrian and modern cyanobacterial look-alikes? What are their names?
Prof. J. W. Schopf: "Precambrian and modern cyanobacterial look-alikes have been known since 1968, when the 'morphologically evolutionary conservation' of the group was first pointed out. Since then, so many fossil-modern look-alikes have turned up that it has become standard practice for the fossils to be named after their living relatives.
"For example, fossilized microbes are (literally) dead ringers (= strongly resemble) for living cyanobacteria of the genus Oscillatoria have been named Oscillatorites ('related to Oscillatoria'), Oscillatoriopsis ('Oscillatoria-like'), and Archaeoscillatoriopsis ('ancient Oscillatoria-like'). To highlight such relations, many workers have simply added the prefixes palaeo- ('old') or eo- ('dawn') to names of living genera. Nearly fifty namesakes have been proposed by workers worldwide for fossil relatives of living cyanobacteria in eight different taxonomic families.
Cyanobacterial fossil namesakes, coined by scientists in Brazil, Canada, China, India, Israel, Russia, USA:
Chroococcaceae Oscillatoriacea
|
Anacystis |
Palaeoanacystis |
Lynbya |
Palaeolynbya |
|
Microcystis |
Palaeomicrocystis |
Spirulina |
Palaeospirulina |
|
Gloeocapsa |
Eogloeocapsa |
Microcoleus |
Eomicrocoleus |
|
Synechococcus |
Eosynechococcus |
Phormidium |
Eophormidium |
|
Aphanocapsa |
Eoaphanocapsa |
Oscillatoria |
Oscillatoriopsis |
|
Eucapsis |
Eucapsamorpha |
Schizothrix |
Schizothropis |
Pleurocapsaceae Nostocaceae
|
Pleurocapsa |
Eopleurocapsa |
Nostoc |
Palaeonostoc |
|
Pleurocapsa |
Palaeopleurocapsa |
Anabaena |
Anabaenidium |
Rivulariaceae Scytonemataceae
|
Calothrix |
Palaeocalothrix |
Plectonema |
Eoplectonema |
|
Rivularia |
Primorivularia |
Scytonema |
Palaeoscytonema |
Entophysalidaceae Hyellaceae
|
Entophysalis |
Eoentophysalis |
Hyella |
Eohyella |
After: J. W. Schopf, Cradle of Life (1999:214, 215).
Have the cyanobacteria changed at least metabolically during those billions of years?
Prof. J. W. Schopf: "The vast majority of microbes that look like cyanobacteria have the oxygen-consuming physiology of full-fledged members of the group. ... From early in biological history to today, the same families, genera, and even species of cyanobacteria have inhabited the same settings, lived in the same kinds of microbial communities, and built the same thinly layered, high-rise stromatolitic condos. This remarkable set of relations could have been sustained only if the metabolic lifestyle of cyanobacteria remained unchanged over the eons." (1999:218).
"In addition to chroococcacean balls and oscillatoriacean strings, three other cyanobacterial families are fairly common in the early fossil record: the Entophysalidacea, Pleurocapsaceae, and Hyellaceae.
"Entophysalidaceans such as Entophysalis and its fossil look-alike Eoentophysalis are composed of jelly-bean-shaped cells that form lumpy, slime-embedded colonies on rocky substrates. Pleurocapsaceans such as Cyanostylon and its fossil counterpart Polybessurus are egg-shaped cyanobacteria that live in close-packed groups where they sit atop long slender gelatinous stalks that radiate upward from the seafloor in pincushion-like lumps. Hyellaceans, represented in modern settings by Hyella and in the early fossil record by Eohyella, are endoliths, cyanobacteria that etch tiny caves in limestone pebbles, boulders, and stony pavements which they then inhabit, living within the outermost rock rind where sunlight penetrates.
"All five cyanobacterial families - the Chroococcaceae (balls), Oscillatoriacea (cellular and tubular strings), Entophysalidacea (jelly beans), Pleurocapsacea (stalked eggs), and Hyellacea (cavity-inhabiting endoliths) - display status quo, hypobradytelic evolution." - (Hypobradytelic = very slowly evolving).
"Hypobradytely is especially well documented for the balls and strings, the most common kind of cyanobacteria in the early fossil record. Comparison of more than 600 species of living cyanobacteria with a worldwide sample of Precambrian fossils, both chroococcacean (nearly 2,000 taxonomic occurrences in about 300 geologic formations) and oscillatoriacean (750 occurrences in 200 formations), shows that practically all of the fossils can be placed in present-day genera, and up to 40% cannot be told apart from particular living species.
"All colony forms known in modern groups are present among the fossils, and the fossil tubular sheaths are identical in shape, size, and detailed structure to those of living counterparts. Most of the several hundred fossil-bearing rock units were laid down in coastal lagoons, mudflats, and tide-washed shallow platforms. From early in the Precambrian to today, these same settings have been inhabited by the same suite of chroococcacean and oscillatoriacean cyanobacteria." (1999:228).
"Status quo evolution is typical of other cyanobacteria as well including distinctive jelly-bean-shaped members of the Entophysalidaceae. One of several excellent examples is the Canadian 2,100-Ma-old genus Eoentophysalis, which is all but identical to modern Entophysalis - in cell shape, colony form, the way the cells divide and grow, the stromatolitic structures they build, the environments they inhabit, the makeup of their microbial communities, even the way their cells break down when they die."
"Egg-shaped pleurocapsaceans also changed little over time. Fossils known as Eopleurocapsa and Paleopleurocapsa are indistinguishable from species of the modern genus Pleurocapsa. And pincushion-like clumps of the stalked, egg-shaped cells of Polybesserus, known from 770-Ma-old stromatolites of South Australia and East Greenland, have the same morphology, reproduction, and pattern of growth as the pleurocapsacean Cyanostylon living today in coastal waters of the Great Bahama Bank, the same environment inhabited by the fossil look-alike.
"Further evidence is found in Eohyella, a Proterozoic hyellacean cyanobacterium known for its unusual rock-boring endolithic lifestyle and described by its discoverers as a 'compelling example of the close resemblance between Proterozoic prokaryotes and their modern counterparts,' a fossil 'morphologically, developmentally, and behaviorally indistinguishable' from living Hyella in the eastern Caribbean.
"Cyanobacterial hypobradytely (= very slowly evolving) is backed by an impressive body of scientific data, the nearly identical organismal form, cellular size and shape, growth characteristics, behavioral traits, and environmental settings shared by hundreds of examples of fossil-modern look-alikes belonging to five very different families. The bottom line is undeniable: cyanobacteria changed little or not at all since they came on the scene billions of years ago.
"This is surprising, even startling. ... Yet during the Precambrian, the most successful of life's early branches evolved at an almost imperceptibly sluggish rate, and for diverse members of this group hypobradytely is the rule, not the exception. Why did cyanobacteria change so little over their exceedingly long history?" (1999:229-231).
"Cyanobacteria are remarkable. They have the longest fossil record, changed least over geologic time, were monarchs of the living world for most of its existence, and invented oxygen-releasing photosynthesis on which later life depends. ... The versatility of cyanobacteria is shown especially by the Chroococcaceae and Oscillatoriaceae, the balls and strings so abundant in the early fossil record. They live, even flourish, in almost total darkness to extreme brightness. In pure, salty, or the most saline waters. In acid hot springs or lakes so alkaline almost nothing else survives. In scalding ponds (70-74°C) or frigid ice fields.
"In the near-absence, presence, or huge overabundance of oxygen or carbon dioxide. In the driest locale on Earth, the Chilean Atacama Desert, where rainfall has never been recorded. Even in the deadly radiation of a thermonuclear blast! ... The success of cyanobacteria comes because they are generalists, able to survive and grow under the most varied conditions. They have no need to evolve, for even if they are outcompeted in a local setting they easily find refuge in places their competitors cannot endure. ... Suited to an amazingly wide range of habitats there was no need for them to ever change." Schopf, J. W. (1999:323-234).
The bacterial cell has not changed at all into any higher form of life. The cell itself has not "discovered" or "invented" anything. The cyanobacterium has remained, what it has been since the beginning, some 3.5-3.8 billion years ago. Because it was perfectly adapted to its environment. It did not need to evolve into anything else. Expressions like "cyanobacterial hypobradytely" (cyanobacteria have evolved very little), and "status quo of evolution", "they evolved at an almost imperceptibly sluggish rate", "morphologically evolutionary conversation", and "hypobradytely is the rule" is just a euphemistic (inoffensive) way of saying to the evolutionistic believer: They have not evolved at all.