Why are there living cells? Why are they able to live? Why is the cell able, to make its own building blocks? How complicated are its chemical reactions? If you, dear reader, do not understand some of the technical terms (or even most of them), do not worry about it. In this way I just want to show you, what the tiny cell all knows about biochemistry, and what it is able to do. Then ask yourself, please: Why does this small one-celled organism know all this about molecular biology and biochemistry, what a professor of molecular biology and biochemistry first had to learn? Many of the technical details within the living cell are so complicated, that the experts themselves have been able to understand now only a few of them. Some they have been able to understand only a little bit, and some, not at all. At least not until now.
Metabolism. Bruce Alberts is an American Professor of Biology. He is President of the National Academy of Sciences (NAS) of America. He and his co-workers do believe that life on earth has evolved by itself. They even claim in their textbook The Cell (1989:41): "There is no room in the contemporary view of life ... for anything else outside the laws of chemistry and physics."
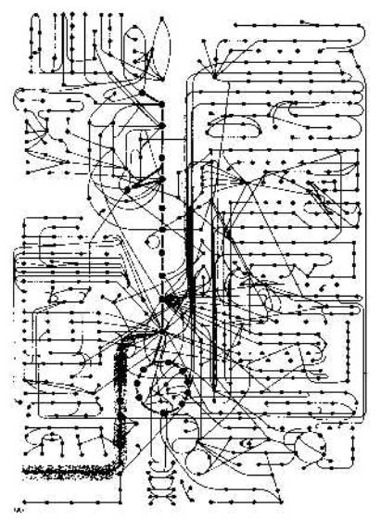
But Professor Bruce Alberts then says: "Some idea of how intricate a cell is when viewed as a chemical machine can be obtained from Figure 2-35 (in his book). It is a chart showing only some of the enzymatic pathways in a cell. All of these reactions occur in a cell that is less than 0.1 mm in diameter. And each requires an enzyme that is itself the product of a whole series of information-transfer and protein-synthesis reactions. For a typical small molecule - the amino acid serine, for example - there are half a dozen or more enzymes that can modify it chemically in different way.
"It can be linked to AMP (adenylated) in preparation for protein synthesis, or degraded to glycine, or converted to pyruvate in peparation for oxidation. It can be acetylated to acetyl CoA or transferred to a fatty acid to make phosphatidyl serine. All of these different pathways compete for the same serine molecule. And similar competitions for thousands of other small molecules go on at the same time. One might think that the whole system would need to be so finely balanced that any minor upset, such as a temporary change in dietary intake, would be disastrous.
"In fact, the cell is amazingly stable. Whenever it is perturbed, the cell reacts so as to restore its initial state. It can adapt and continue to function during starvation or disease. Mutations of many kinds can eliminate particular reaction pathways, and yet - provided that certain minimum requirements are met - the cell survives. It does so because an elaborate network of control mechanisms regulates and coordinates the rate of its reactions. ... Here we are concerned only with the simplest mechanisms that regulate the flow of small molecules through the various metabolic pathways." Alberts, B. et al. (1989:80).
Metabolic Pathways regulated. The metabolic pathways in the cell are regulated. How?
Prof. Bruce Alberts: "The concentrations of the various small molecules in a cell are buffered against major changes by a process known as feedback regulation, which fine-tunes the flux of metabolites through a particular pathway by temporarily increasing or decreasing the activity of crucial enzymes. For example, the first enzyme of a series of reactions is usually inhibited by a negative feedback effect of the final product of that pathway: if large quantities of the final product accumulate, further entry of precursors into the reaction pathway is automatically inhibited. Where pathways branch or intersect, as they often do, there are usually multiple points of control of different final products. The complexity of such feedback control processes is illustrated in Figure 2-37 (in his book). It shows the pattern of enzyme regulation observed in a set of related amino acid pathways.
"Feedback regulation can work almost instantaneously and is reversible; in addition, a given end product may activate enzymes leading along other pathways, as well as inhibit enzymes that cause its own synthesis." (1989:80).
The coordination of catabolism (breaking down) and biosynthesis (making new) in the cell. After B. Alberts et al. (1989). The biochemical circuit diagram has been thought out by an intelligent person: the Creator of heaven and earth. Man has only found and understood it a little.
How does the bacterial cell control its fueling reactions? Why does it know, which fueling reaction is right and which one is wrong?
Prof. F. C. Neidhardt and co-workers: "Allosteric inhibition and activation play important roles in regulating the flow through fueling pathways. Here the simple device of end product control of the first or an early step in its formation cannot apply. The pathway of formation of the 12 precursor metabolites are far too interrelated (some are cyclic) for each to control its own formation by inhibiting a unique early step - there are no unique early steps.
"Instead, controls work internally in each of the main fueling pathways (Figure 4 in his book). For example, phosphoenolypyruvate, an intermediate of glycolysis, inhibits phosphofructokinase, an enzyme catalyzing a reaction three steps back in the same pathway. And a-ketoglutarate, an intermediate of the tricarboxylic acid cycle (TCA), inhibits citrate synthase, which catalyzes the reaction two steps back.
"It should be remembered that not all the control on central fueling reactions can be accomplished by allosteric interactions. The reversal of flow in the Embden-Meyerhof-Parnas (EMP) pathway to accomplish gluconeogenesis during growth of cells on malate or pyruvate, for example, can occur because all but two of the reactions between pyruvate and hexose phosphate have equilibrium constants not far from 1. And the two that are practically irreversible are bypassed by two other enzymes - phosphenolpyruvatesynthetase, which bypasses pyruvate kinase, and a specific phosphatase that hydrolzyes fructose 1,6-diphosphate.
"Balancing the rates of formation of the 12 precursor metabolites to one another and to the demands of the biosynthetic pathways involved the action of these compounds as allosteric effectors, as illustrated partially in Figure 4. But the need for coordination goes beyond the carbon-containing metabolites. Fueling reactions must supply ATP and reduced pyridine nucleotides (both NADH2 and NADPH2). It is not surprising, therefore, to find that these compounds are allosteric effectors at several points in the central fueling pathways." (1990:310, 311).
When eating, when repairing itself, and when doubling itself, the one-celled organism must do many things at the same time and in the right order. How does this tiny creature do that?
Prof. F. C. Neidhardt and co-workers state in their textbook Physiology of the Bacterial Cell under the heading "Regulation of Gene Expression: Multigene Systems and Global Regulation":
"Many bacterial activities involve gene coordination at levels of organization higher than individual transcriptional units. Most, if not all, the operons of a bacterial cell belong to higher level regulatory organizations, which we shall call REGULATORY NETWORKS:
"The operon is a hallmark of the procaryotic cell. This mode of organization - whereby the genes of an entire pathway can be united as a single transcriptional unit - achieves a simple solution to the problem of coregulating genes of related function. Why go further? Is there a need for the bacterial cell to move beyond this obvious successful strategy?
"There are at least two answers to these questions. The first is that some bacterial processes involve too many genes to be accommodated in a single workable operon. This situation is illustrated by the translation machinery, a group of at least 150 gene products (ribosomal RNA, ribosomal proteins, initiation, elongation, and termination factors, amino-acyl-tRNA synthetases, and tRNAs) involved directly in making protein from amino acids.
"They are so numerous that coordinating their synthesis by linking their genes into a single operon would be awkward, if not impossible. Yet, coordinated regulation of these genes is important to the overall efficiency of bacterial growth. ... This coordination is accomplished even though these genes are organized into many dozens of unlinked operons.
"The second answer is that some bacterial processes involve a number of genes that must be both independently regulated and subject to an overriding, coordinating control. This situation is illustrated most clearly by the ensemble of genes encoding catabolic enzymes involved in the utilization of sugars, amino acids, and other compounds for carbon and energy. During growth in an environment containing a mixture of such compounds, economy would demand that only a premium substrate, best able to satisfy the carbon and energy needs of the cell, be metabolized. (For enteric bacteria, glucose is such a substrate; for pseudomonads, succinate is.) The operons encoding enzymes for the metabolism of the secondary, redundant substrates should be repressed. Each operon must, however, be induced individually when its cognate substrate is present in the absence of the premium substrate. This double requirement calls for a level of organization above that of the operon." (1990:351, 352).
How many multigene systems are there in a bacterial cell? How do they work? And how complex are they?
Prof. F. C. Neidhardt and co-workers: "It is estimated that the bacterial cell has evolved several hundred multigene systems. Their discovery and analysis has only just begun. Some of the better-known examples for the enteric bacterium Escherichia coli are:
· Networks involved in response to limitation of one or another nutrient - carbon and energy sources, ammonia, inorganic phosphate
· Networks involved in oxidation-reduction reactions and electron transport
· Networks concerned with response to damage by oxidation, radiation, high and low temperature, and extremes of osmotic pressure
· Miscellaneous networks, some with subnetworks, that bring about major changes in the physiology and morphology of the cell.
The bacterial cell has evolved diverse ways to weave individual operons into coordinated networks; the various mechanisms are just beginning to be elucidated. In some cases the devise of an allosteric protein regulator has simply been borrowed from operon regulation: a protein repressor or activator recognizes a particular sequence common to the controlling regions of the member operons. This devise is used in the SOS, oxidation damage, and anaerobic electron transport systems in enteric bacteria.
"In other systems the network is defined by an alternative sigma factor that reprograms RNA polymerase to recognize the promoters of the member operons. The heat shock and sporulation systems of various bacterial species illustrate this situation. Other networks involve a combination of protein regulators and sigma factors, as found in the nitrogen utilization system of many bacteria. One of the most pervasive networks, the stringent operons are regulated by the nucleotide guanosine tetraphosphate (ppGpp) in a manner still to be elucidated.
"Because many regulatory networks were first recognized as cellular responses to environmental changes, the formal structure of a stimulus-response system can provide a useful organizing function for the mass of information about each network. A stimulus-response system (Figure 1 in his book) contains a pathway that begins with a stimulus from the environment that affects some cellular target, or sensor, which generates a signal. This stimulus-response system directly - or indirectly when the signal passes through one or more transducers - affects the activity or synthesis of a regulator, which controls the output, usually some adaptive response to the environmental change. This system often includes a feedback control mechanism that permits a return to the prestimulus condition or to a new equilibrium consonant with a changed environment.
"In a bacterial gene network, a stimulus from the environment (for example, changed temperature, nutrient status, or toxicity) would be depicted as affecting some sensor which would generate a signal to affect the activity or synthesis of (usually) a protein regulator of the member genes. The products of the member genes would perform some cellular function(s) to promote the growth or survival of the cell. Modulators of some sort would enable a return to the prestimulus state of expression of the member operons or would establish a new level of their expression.
"It is clear ... that many of the operons of E. coli are already assigned to one or more networks. With the help of the framework provided in Figure 1, we shall prove a few of these networks in depth to illustrate principles of multigene physiology and molecular biology. The first two networks to be discussed have been chosen partly because of their quantitative importance in cell physiology; together they directly or indirectly control probably three-fourth of the protein-synthesizing capacity of the bacterial cell." Neidhardt, F. C. (1990:352-357).
Catabolite repression of fueling genes: what does that mean? And how does that work?
Prof. F. C. Neidhardt and co-workers: "Most species of bacteria posses an extensive repertoire of genes encoding catabolic enzymes directed against dozens of different compounds of diverse nutritional value. Growth on a single source of carbon and energy - a substrate - requires a relatively high cellular level of the enzymes that metabolize the substrate and feed the catabolic products into the central fueling pathways, because all metabolic pathways in the cell flow from the metabolites produced by these catabolic enzymes.
"Bacteria appear to sense the appropriateness of each catabolic pathway in a given circumstance and to regulate gene flow expression accordingly. We have already noted ... that the program for deciding what enzymes to make and in what quantities most be more sophisticated than one, which simply calls for the cell to induce catabolic enzymes for substrates sensed in the environment. Indeed, despite a half-century of intensive effort, the sophistication of this cellular program has exceeded the ability of physiologists to understand its intricacies.
"Enteric bacteria employ at least four different processes to assure the priority status of glucose as a carbon and energy source:
· Constitutive synthesis: The enzymes of glucose catabolism, being in large measure a part of the EMP central fueling pathway, are produced at high levels no matter what substrates are present in the environment.
· Inducer exclusion: Glucose prevents the entry of certain other inducing substrates by inactivation of their permeases.
· Transient repression: Addition of glucose produces a transient, severe inhibition of synthesis of inducible catabolic enzymes for approximately half of a generation despite the presence of their inducer.
· Catabolic repression: Metabolism of glucose produces a continued, but less severe inhibition of induced enzyme synthesis, even in the presence of the appropriate inducers.
"Non of these processes are fully understood, and catabolite repression is particularly complex. It includes one element, however, about which much has been learned: the CAMP-CAP NETWORK. In E. coli the lac operon is a member of this network. The total number of member operons is not known; but it may be several dozen and includes those concerned with metabolism of galactose, arabinose, maltose, tryptophan, D-serine, and histidine. Member operons possess a characteristic nucleotide sequence to which the regulatory protein CAP (catabolic gene activator protein, product of the crp gene) binds." Neidhardt, F. C. et al. (1990:357, 358).
Signal transduction in multigene systems, what does that mean? How does that work? And how complex is it?
Prof. F. C. Neidhardt and co-workers: "The arc network with the other three networks just described, share a common theme: information processing by protein-protein interaction. Let us consider this topic in greater depth.
"The processing of physiological information is not a new topic for us. We encountered some aspects of information processing when we considered the conformational changes in certain (allosteric) proteins brought about by reversible complexing of specific small molecules. The small molecules - end products of biosynthetic pathways, inducers of catabolic pathways, precursor metabolites, or nucleotides - conveyed information (about their cellular concentration) to the allosteric protein. This process accounted for most of the control on enzyme activity in biosynthetic pathways and in fueling reactions, and for the central strategic device of operon physiology - activation and repression of transcription by proteins that in one conformation can bind DNA and in another, cannot." (1990:375-377).
How does the bacterial cell regulate its multigene networks? Why are these interactions important?
Prof. F. C. Neidhardt and co-workers: "Protein-protein interactions are important in regulating the networks described above but are not the sole means of regulating multigene networks. In fact, the molecular means by which groups of different operons are coordinated in their expression are quite diverse.
"To illustrate this diversity, we consider three networks that resemble one another in one functional aspect. Each is induced to high-level expression as the result of a single, well-defined environmental parameter change, but the mechanism of the induction is very different in each instance.
"A shift to high temperature induces a set of approximately 20 genes in most bacterial cells. This induction constitutes the heat-shock response, which, to an incredible extent, is conserved from archaebacteria and eubacteria up through lower eucaryotes to higher plants and animals. Virtually every microbial cell and every plant and animal cell responds to a shift-up in temperature to a range just above that which is normal for the given organism by inducing the synthesis of its heat-shock proteins.
"Another network - with approximately the same number of genes - is the SOS system. This group of genes is induced to high-level expression when the cell experiences damage to is DNA, as by UV irradiation. The protein products of these genes are involved in various processes that repair damage to DNA and that prevent the cells from dividing until such damage is repaired. This network is induced by a proteolytic cleavage of the repressor of the member operons.
"The repressor is the product of the lexA gene; upon damage to the DNA of a cell, the RecA protein is activated to facilitate the proteolytic cleavage of LexA, thereby inducing the LexA-controlled genes, which constitute the SOS system.
"Still another mode is illustrated by the oxyR network. This group of a dozen or so genes is induced by oxidative damage brought on by, for example, hydrogen peroxide. The member genes encode protective enzymes such as catalase-hydroperoxidase I and alkylhydroxyperoxidase. This network is induced by activation of the positive regulator OxyR, product of the oxyR gene.
"These examples serve to illustrate the fact that the regulation of gene networks is as diverse as is the regulation of individual operons." Neidhardt, F. C. (1990:380, 381).
Why does the living cell make proteins? And why does it know, how to make them? And why does a certain set of nucleic acids translate the text into proteins? In other words: Why does a certain sign (or letter) of the DNA code know, what it means? And why does it mean this?
Bruce Alberts and co-workers do admit in their textbook Molecular Biology of the Cell (1989:219), though they do believe in evolution: "The molecular processes underlying protein synthesis seem inexplicably complex. Although we can describe many of them, they do not make conceptual sense in the way that DNA transcription, DNA repair, and DNA replication do. As we have seen, protein synthesis in present-day organisms centers on a very large ribonucleoprotein machine, the ribosome. It consists of proteins arranged around a core of rRNA molecules.
"Why should rRNA molecules exist at all? And how did they come to play such a dominant part in the structure and function of the ribosome? The answer would undoubtedly help us to understand protein synthesis. ... Protein synthesis also relies heavily on a large number of different proteins that are bound to the rRNAs in a ribosome. The complexity of a process with so many different interacting components has made many biologists despair of ever understanding the pathway by which protein synthesis evolved."
We still might remember, what Professor Bruce Alberts and his co-workers claimed in their textbook The Cell (1989:41): "There is no room in the contemporary view of life ... for anything else outside the laws of chemistry and physics."
Is that logical, is that reasonable, is that scientific? Not at all. The statement: "There is no room in the contemporary view of life for anything outside the laws of chemistry and physics" is not logical, it is not reasonable, it is not scientific. The reason: A law always comes from a lawgiver. The laws of chemistry and physics do contain highest scientific information and mathematics. Human scientists down here on earth have understood until now only a few of them. Some of these laws of chemistry and physics they have been able to understand only a little bit. Information and mathematics do exist independently of mankind. The human scientist has only found and understood them a little. Information and mathematics are something non-material, spiritual. They have their source in a spiritual, non-material world, in the true God of the Bible, whose name is Jehovah.
Already on the level of the code we can know, that it has been thought out and made by an intelligent person. A code (or alphabet) is something spiritual, non-material. An intelligent person must first think it out. He must first select certain signs (letters, etc.) and decide, what they shall mean. With these signs he can write then the words, sentences, chapters, and the whole book.
· The 20 amino acids are the 20 letters of the amino acid code. With these 20 letters (and the grammar supplied with it), the living cell makes (writes) its proteins.
· The 4 nucleic acids are the 4 letters of the nucleic acid code. With these 4 letters (and the grammar supplied with it), the living cell makes (writes) its genetic code, its DNA chain.
The first archaeal cell on earth was just as complex as those, living now. Even the most "simple" one-celled organism is more complex, than anything, which man has be able to make till now. The fastest super-computer of today is just a primitive child's toy, compared to the simplest cell, which is living today, and which has lived at the beginning on this earth, some 3.8-4.0 billion years ago.
The hypothesis of evolution, or neo-Darwinism, as now commonly taught throughout the world, has nothing to do with serious science at all. It is a false religion. It has its roots in the teachings of ancient Babylon (Sumer-Accad) and Egypt. Their pagan priests taught already some 3.500 to 4000 years ago, that life on earth has evolved by itself from the water and mud of the Euphrates and Tigris and from the Nile. The modern hypothesis of evolution or neo-Darwinism is a huge fraud, hiding its true meaning behind the white dress of modern science.