Gram-positive and Gram-negative bacteria. Their membrane and wall. How they make their walls. How they differ. What was needed, to think them out and to make them? Does their cell wall prove that they all have come from a common ancestor?
Prof. Frederic C. Neidhardt and co-workers write in their textbook Physiology of the Bacterial Cell (1990:25-27) about the wall of the bacteria: "Bacteria can be divided into two broad classes - Gram-positive and Gram-negative... Gram-positive and Gram-negative bacteria differ in a number of important aspects in addition to envelope structure. ... The Gram-positive-Gram-negative distinction is a profound one. It reflects not only a fundamental difference in wall structure but other important biochemical, physiological, and genetic differences as well. The majority, but not all, bacteria, are members of these two groups. Those that fall outside these groups are of two classes. One group is closely similar to other bacteria, but completely lack walls - the Mollicutes. The other group, the Archaebacteria, produce walls that do not contain a typical murein, and in many other respects differ from the typical procaryotic cell." (1990:25-27).
What have scientists found out now about the cell wall? How complex is it? What must one all know, in order to understand and to make it?
Prof. F. C. Neidhardt and co-workers: "As cells go, bacteria are particularly tough; that is, they are hard to break by mechanical means, and they retain their shape even under harsh conditions. Much of the toughness of bacteria is related to their cell wall, which provides rigid mechanical support and prevents pressure from bursting the cells (osmotic lysis). In addition, the cell wall represents a chemical and physical defense against noxious chemicals that may harm the cell membrane. Gram-positive and Gram-negative bacteria differ considerable in the structure of their cell walls and in the role of this structure in coping with environmental changes (Figure 12 in their book). Any portion of the wall of Gram-positive cells looks like a thick blanket, that of most Gram-negative cells like a flimsy sheet. In reality, even the thin Gram-negative wall has considerable tensile strength." (1990:33).
Prof. F. C. Neidhardt and co-workers: "The Gram-positive cell wall consists of a thick multimolecular coat of muerein (a type of peptidoglycan) with lesser amounts of other polymers, notably teichoic acids; interspersed. The polymeric fabric consists of many layers wrapped around the length and width of the cell, thereby forming a sac that determines the size and shape of the organism. The murein can be isolated intact as a structure called the murein sacculus. The shape of the bacterium depends on the shape of the sacculus: most bacteria look like rods (bacilli), or helixes (spirochetes); a few look like spindles, starfish, or flattened polyhedra; still others assume more complex, differentiated shapes. ... The rigid murein meshwork allows bacteria to survive in media that are commonly hypotonic - having a lower osmotic pressure than that of their cytoplasma. In the absence of a rigid corset-like structure to push against, the membrane would burst and the cells lyse (= flow apart)." (1990:33-36).
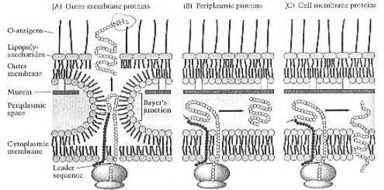
Prof. F. C. Neidhardt and co-workers: "Gram-negative bacteria have evolved a radically different solution to the problem of protecting the cytoplasmic membrane. Their murein layer is much thinner than that of Gram-positive bacteria, and they make a completely different structure - an outer membrane, which is built up outside a thin murein layer. The outer membrane is chemically distinct from the usual biological membranes and has the ability to resist damaging chemicals. It is a bilayered structure, and its inner leaflet resembles in composition that of the cytoplasmic membrane.
"Its outer leaflet, on the other hand, has a unique constituent in the place of phospholipids. This component is the bacterial lipopolysaccharide, or LPS, a complex molecule not found elsewhere in nature. As a result, the leaflets of this membrane are extremely asymmetrical, and the properties of this unusual bilayer differ considerably from those of a regular biological membrane. The ability to exclude hydrophobic compounds is unusual among biological membranes.
"Exclusion of hydrophobic compounds in Gram-negative bacteria, as in Gram-positive bacteria, is accomplished by surrounding the cells with hydrophobic polysaccharides, but these differ in structure and organization in the two groups. The outer membrane, however, presents an apparent impasse: because of its lipid nature, it can be expected to exclude hydrophilic compounds as well. In that case, no compounds, hydrophobic or hydrophilic, could cross the outer membrane.
"While solving the problem of protection of the cytoplasmic membrane, Gram-negative bacteria appear to have created a new one. How do these organisms transport their nutrients? Do they copy in the outer membrane the active transport devices of the cytoplasmic membrane? This strategy not only would be a wasteful investment, but also could make the outer membrane just as sensitive to environmental challenges as the cytoplasmic membrane is. Once again, bacteria have found an interesting solution: the outer membrane has special channels that permit the passive diffusion of hydrophilic compounds like sugars, amino acids, and certain ions. These channels consist of protein molecules, aptly called porins. Porin channels are narrow, just the right size to permit the entry of compounds smaller than 600-700 daltons (Figure 9 in their book).
"The channels are small enough that hydrophobic compounds would come in contact with the polar 'wall' of the channel and thereby be excluded. To some extent, E. coli can select porins with different channel sizes in response to the osmotic properties of the medium.
"Certain hydrophilic compounds necessary for the cell's survival are larger than the exclusion of porins. These large molecules include vitamin B12, sugars bigger than trisaccharides, and iron chelates. Such compounds cross the outer membrane by separate, specific permeation mechanisms, which utilize proteins especially designed to translocate each of these compounds. Thus, the outer membrane allows the passage of small hydrophilic compounds, excludes hydrophobic compounds, large or small, and allows the entry of some larger hydrophilic molecules by especially designed mechanisms." (1990:37-40).
"The outer membrane is not an entirely separate structure; it is connected to both the murein layer and the cytoplasmic membrane. The connection with the murein layer is mediated by two types of interactions, the most important being that involving an outer membrane lipoprotein. This protein is present in some 700,000 copies per cell, which makes it the most abundant protein (numerically, not by weight) in E. coli.
"About one-third of the lipoprotein molecules are covalently linked to murein and help hold the two structures together. The other type of interaction is the tight (but probably not covalent) association of some of the outer membrane porins with murein. Together, lipoproteins and porins provide in each cell over 400,000 strutlike contacts between the outer membrane and the cell wall." (1990:41).
Which specific substrates is the bacterium E. coli able to take up? How does it take them up, in which different ways?
Prof. F. C. Neidhardt and co-workers: "Of the various transport mechanisms, almost all (including facilitated diffusion, shock-sensitive active transport, and secondary active transport) participate in the uptake of one or another sugar or sugar alcohol by E. coli (Figure 4 in their book). Interestingly, the uptake of a particular sugar does not follow a pattern: for example lactose is taken into E. coli by proton transport, but into Staphylococcus aureus by a PTS.
"Amino acids are transported by shock-sensitive or secondary active transport systems. In E. coli, 14 different transport systems are dedicated to bringing amino acids into the cell. From an examination of the amino acid transport systems of bacteria, several patterns emerge. Some of the systems transport a group of amino acids with similar structure. Often these have subsystems for only one of the amino acids. This apparent redundancy serves a purpose. One of the systems has high affinity (affinity constant 10 times or more greater) and high flow. Each has obvious advantages in particular environments. In addition to amino acid transport systems, E. coli has a variety of transport systems for cofactors, various ions, and metabolic intermediates." (1990:182, 183).
How a Gram-negative-bacterium makes its own envelope proteins. From Neidhardt et al. (1990:114) Fig. 8. The tiny cell has achieved here, what man is still not able to do at the beginning of this 21st century: automatic assembly, automatic production. So the One, who has thought out and made this one-celled organism, must know much more about science and technology, than any scientist now living on this Earth: the Creator, whose name is Jehovah.
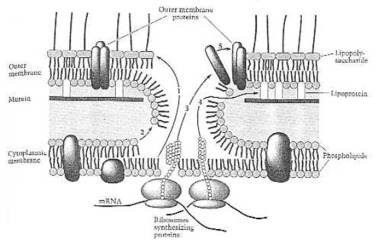
Assembly reaction of the outer membrane. (1) Lipopolysaccharide;(2) phospholipids; (3) outer membrane proteins; (4) lipoprotein; (5) self-assembly of the outer membrane. From Neidhardt et al. (1990:121) Fig. 14.
How does E. coli overcome the extremes of environmental osmotic pressure? And how does it make its porins, the pores in its outer membrane?
Prof. F. C. Neidhardt and co-workers: "One important environmental variable with which bacteria must deal is the osmolarity of their environment. The bacterial cell utilizes several molecular strategies to counteract the effects of deleterious extremes of environmental osmotic pressure.
"The outer membrane is traversed by channels lined by proteins called porins. In E. coli there are two major porins - OmpF and OmpC. They tend to be made as alternatives to each other. The pores made by OmpC are slightly smaller and are made preferentially at higher osmotic pressure and higher temperature. This pattern means that OmpC is the predominant form during growth of enteric bacteria in the intestinal tract. It makes good physiological sense for enteric bacteria to have smaller pores when residing in the intestinal tract and larger ones outside. The smaller pores exclude many of the toxic molecules found in the intestinal tract. The larger pores allow substrates from dilute solution to enter the cell pore readily, because solutes must diffuse through the outer membrane pores and the rate of diffusion is proportional to the cross-sectional area of the pore as well as the concentration difference across the membrane.
"The genes encoding the major porins, as well as certain other genes affecting outer membrane properties, are controlled by two proteins - products of the linked envZ (for envelope proteins) and ompR (for outer membrane protein regulation) genes. The EnvZ protein is a cytoplasmic membrane protein that is thought to function as an osmo-sensor. It has strong structural similarity to certain protein kinases; on the basis of this similarity and other evidence, and from the phenotype of missense and other mutations in envZ, investigators believe that EnvZ interacts with the second protein, OmpR, affecting its activity by phosphorylating it.
"Protein OmpR binds specifically to the DNA upstream of promoters that it controls; the binding site is around -50 in the ompF promoter, and -90 in the ompC promoter. One current model for how alternative expression of ompF is accomplished is as follows (Figure 7 in their book): high osmotic pressure leads to phosphorilation of EnvZ, and this activated protein in turn phosphorylates OmpR. Phospho-OmpR (OmpR-P) represses transcription of ompF by binding to a site between -40 and -60 in the ompF promoter region; it activates ompC transcription by binding to low-affinity sites between -75 and -105 in the ompC promoter region. When the osmotic pressure is low, the level of OmpR-P is low, and therefore it is unable to activate ompC; but it is still able to activate ompF by binding to high-affinity sites in the -60 to -100 region of the ompF promoter.
"The effects of OmpR-P as a stimulator and as inhibitor of transcription of these two genes is supplemented by another mechanism. The gene micF is transcribed in the opposite direction from ompC from a promoter near that of ompC; micF encodes a 174-base RNA that is complementary to a region near the start of the ompF message. The micF transcript has been shown to block expression of ompC. Because the regulation of micF expression is similar to that of ompC, the conditions that induce ompC will simultaneously induce micF, and thereby help repress ompF (Figure 7 in their book).
"There is a network of genes besides those for the two porins that are regulated by the envZ/OmpR pair of interacting proteins; some of these genes are induced, others repressed. The overall physiological sense of the choice of genes that are included in this system is still elusive; the importance to us of the EnV/OmpR story is that it has provided an important pattern of protein-protein interaction in stimulus-response networks, including many for which the physiological function is crystal clear." Neidhardt, R. C. et al. (1990:272, 373).
Secondary active transport systems. (A) Symport reactions. The pH gradient created by primary active transport drives (on the left) an electroneutral symport of an anion with a proton, and (on the right) an electrogenic symport of an uncharged solute (S) together with a proton. (B) Antiport reaction. The pH gradient drives (on the left the electroneutral exchange of a cation for a proton and (on the right) the electronic exchange of an uncharged solute (S) for a proton. (C) Uniport reactions. The pH gradient drives a cation into the call (left) of an anion out (right). From Neidhardt et al. (1990:178) Fig. 1.
![]()
Carrier-mediated mechanisms for the uptake of sugars and sugar alcohols by E. coly. Glycerol (Glyc) enters by facilitated diffusion. Maltose (Mal) enters by a shock-sensitive system dependent on a binding protein (sphere), two transmembrane proteins (ovals), and an energy-transducing protein (curved, double-headed arrow) that hydrolyzes a high-energy phosphate donor (D~P). Lactose (Lc) enters by protein symport and melibiose (Mel) by a Na+ symport of secondary active transport. Glucose (Glc) enters by a PTS through the mediation of enzymes I (I), HPr, enzyme II (II), and enzyme III (III); the intracellular product of the process is glucose 6-phosphate (G-6-P). From Neidhardt et al. (1990:182) Fig.4.
How has the first membrane come into being? Could it have arisen by itself in the primordial chemical soup?
Prof. Siegried Scherer: "Phospholipides are a main-component of the biomembranes. They are made from glycerin, phosphoric acid and long-chain aliphatic compounds. They are joined together with glycerine as ethers or esters. These molecules, like all other substances, which are active at the surface (tenside) do have an hydrophobic (= water-avoiding, and an hydrophilic (= water-loving, component. Molecules with these structure-characteristics are able, to come together spontaneously. For example, as double-layers, micells, and vesicles). How such substances could have arisen under prebiotic conditions, is not known.
"Biologically active membranes, as we know them from living cells, always originate from already existing membranes. They are not synthesized de novo. The biomembrane must not only demarcate the cell against its surroundings. But it must also, among other things, transport material through its membrane. The complex construction of biomembranes mirrors the diverse tasks of the cell membranes. For example, mechanism of recognition, cell-cell-communication." (1998:146).
Hans Kleinig is a professor for cell-biology at the University of Freiburg. Uwe Maier is a professor for cell-biology at the Philipps-University Marburg, Germany. They write in their textbook Cell-biology (1999:73) about the biosynthesis of membranes: "Biomembranes cannot arise de novo in the Cytoplasma of a cell. The membrane-components, proteins, like lipids, are usually synthesised in the membranes themselves: in protocytes: in the plasma-membrane..."
Biologically active membranes, as we know them from living cells, always originate from already existing membranes. They are not synthesized de novo. How such substances could have arisen under prebiotic conditions, is not known.
We have looked now briefly at Gram-positive cells and Gram-negative cells. The Gram-positive cell has a wall. The Gram-negative cell has a membrane. Gram-positive cells, Gram-negative cells, and archaeabacteria differ much from each other. They are not related to each other at all. They are just as different from each other, as they are from plants and animals. And one cannot trace them back to a mythical common ancestor. Already some 3.5 to 3.8 billion years ago, near the beginning of earth's history, there were Gram-positive and Gram-negative cells, and archaea, just like today.
Many of the genera and species, identified at Warrawoona, NW Australia, are still living today. These one-celled organisms were just as complex, as those living today. They have not evolved at all into higher forms of life. They are fully automatic chemical factories, with a diameter of a tiny fraction of a millimeter. How do chemical factories, houses, with their walls and roofs, arise today? A house (or chemical factory) arises at first in the mind of an intelligent human being.
· First, he has a need. He might say to himself: "I want to have it warm and dry, where the bears and wolves cannot come and kill me."
· Then he has a wish: "I want to have a house (or a tent, or a cave).
· Then he makes a plan: "My house shall be so and so large. It shall have so and so many rooms, doors, and windows. And I shall make it with bricks (or stones, or logs)."
· Then he uses his construction plan (blueprint), energy, and building materials, and makes the house.
The first three steps, leading to the house, are purely spiritual, non-material. An intelligent human being, trained as a house builder, has first thought them out. Even the bricks in the house (their form and function), an intelligent human being has first thought out. The atoms, which make up the bricks (stones and logs) do not know anything about a house.
Materialists have claimed: Everything comes from matter. But a house does not grow up out of the stones or logs of the Siberian taiga, like a tree does. Why not? Because the building material of the house does not know anything about the form and function of a house. All that comes first from an intelligent human being, who has thought about this.
The atoms of the bacterium's wall (or membrane) are at its lowest organizational level. They are only a means to an end, like the bricks in the house. A bacterium is much more complicated, than any house, which man has ever built. Man is able to build a house (and a chemical factory). But he is not able to make living cell, a fully automated chemical factory. It is too complicated. It is more complicated, than anything, which man has ever made till now. The first living cells on earth, some 3.5-4 billion years ago, were just as complicated, as those living today.
The cell contains high biochemical know-how. It contains information and mathematics. But the cell itself has not "discovered" or "invented" anything. Information and mathematics are something non-material, spiritual. They have their source in a spiritual, non-material world. Hence, also the bacterium must first have been thought out and made by someone, who knows much more about science, than any human being, now living on this earth: the Creator, the God of the Bible, whose name is Jehovah.