Also some archaea, living in boiling water, do have a rotary motor with a propeller. The flagellum, driven by a rotary motor, is the world's smallest rotary motor. - Why has it arisen? How does it work? And what was needed, to think it out and to make it? What have scientists found out now about this?
Professor F. C. Neidhardt and co-workers state in their textbook Physiology of the Bacterial Cell, about the bacterium's rotary motor and its propeller: "The flagellum (plural: flagella) is the organ of bacterial locomotion. It is a helical filament that is driven by a motor at its base and rotates relative to the bacterial surface, therefore propelling the cell through the medium. Bacterial flagella impart motion by rotation only, not by bending, as in the case with eucaryotic flagella (thus, for bacteria, the term flagellum, which means 'whip,' is misleading). Flagella are a rare example of rotating shafts in biology, their presence allowing one to come to the conclusion that biological wheels do indeed exist.
"Not all bacteria are motile, and some motile bacteria do not possess flagella. Other forms of motility exist, as seen in the so-called gliding bacteria. Flagellated bacteria can be differentiated by the number and position of these organelles. Some species, such as members of the genus Pseudomonas, have a single, polar flagellum. Escherichia coli has approximately 10 flagella; its relative Proteus, several hundreds. In these species, flagella are inserted all over the surface; such cells are called peritrichous ('hairy all over').
"The flagellum is composed of three parts, which have different molecular complexity. Outermost is the long helical filament, which extends 5 to 10 µm into the medium - several times the length of the cell. The filament is connected via a hook to the basal body, a complex structure that anchors the flagellum to the cell envelope and serves as the motor that turns the flagellum." (1990:47).
Of what is the filament made?
Prof. F.C. Neidhardt and co-workers: "The filament is composed of a protein, flagellin; there are several thousand copies of this protein in each flagellum. In a few species (e.g., Caulobacter), flagella are composed of two types of flagellin, but in most only a single type is found. ... The diameter of a flagellum is about 20 nm in E. coli and is constant throughout its length. The wavelength of the helical turns varies among species but is typically 2 to 2.5 µm per turn.
"Flagellin molecules aggregate spontaneously to form the characteristic structure of the flagellar filament. Isolated filaments can be dissociated into a solution of flagellin in a primer, the flagellin molecules spontaneously reaggregate to form filaments indistinguishable from the natural product. This event is a fine example of biological morphogenis taking place by the self-assembly of molecules. In vitro, the rate of elongation is constant, and 'growth' of the filament is from the end that would be distal to the cell.
"The hook is a short curved structure that connects the flagella filament to the cell. It appears to act as the universal joint between the motor in the basal structure and the filament. The molecular structure of the hook is also quite simple. Like the filament, it is an aggregation of a single type of protein called hook protein. The hook is slightly larger in diameter than the filament and has a constant length - approximately 80 nm in E. coli.
"The flagellar hook is connected to the basal body, which is a small but complex structure embedded in the cell surface. The basal body is composed of 15 or more proteins that aggregate to form a rod to which four rings are attached (at least in Gram-negative cells). The rings appear to act as bushings or as 'stators,' on the one hand anchoring the structure in the various layers of the cell envelope, on the other hand allowing the rod (the 'rotor') to rotate. It is not yet known how the rod portion of the basal body is physically retained on the cell surface.
"As might be expected, the variation in envelope structure between Gram-positive and Gram-negative bacteria is reflected in a difference in the basal bodies of their flagella. Gram-positive cells have only two rings - one embedded in the cell membrane and another associated with the teichoic acid component of the wall." Neidhardt, F. C. (1990:47-49).
Why does the flagellum rotate? What is really behind this?
Prof. F. C. Neidhardt and co-workers: "No one really knows the details but the energy source for the rotation of the basal body and its attached hook and filament is known to be the protonmotive force, or membrane potential. This energy is generated by the electron transport chain during respiration or, anaerobically, by hydrolysis of ATP. The flagellar motor is a highly efficient device and requires the passage of only about 1,000 protons per turn.
"Flagellar motors are a part of a complex behavioral system that enables motile bacteria to move toward environments favorable for growth and away from hostile environments (Macnab, 1987b). How is this done? Flagellar rotation can take place in either direction clockwise or counterclockwise. The choice of direction makes a lot of difference in the outcome: counterclockwise rotation propels the bacterium smoothly forward, a process called swimming. The reason for this difference is that the flagellar filaments are normally left-handed helixes. Thus, rotation in the counterclockwise direction exerts a pushing motion. ... Counterclockwise rotation allows the flagellar filaments to sweep around the cell and to make a common bundle that can operate in concert. Clockwise rotation caused the bundle to disperse.
"Swimming is normally interrupted by episodes of tumbling, and the length of time for each episode is determined by compounds in the environment. The flagellar motor responds to chemical stimuli called attractants or repellants, or, more accurately, to gradients in concentration of such compounds. The concentration of an attractant or repellant determines the length of time flagella will turn counterclockwise or clockwise. The net result of this behavior is chemotaxis, the ability of motile bacteria to swim toward attractants and away from repellants." (1990:49-51).
How does the motile bacterium make its flagellum? What must it all know and be able to do? And how does it assemble then the different parts? Why is the bacterium able to make its own rotary motor and propeller?
Prof. F. C. Neidhardt and co-workers: "Flagella are complex structures composed of a basal body, a hook, and a long filament. They are assembled by a series of intricate steps. At least 40 genes are required for flagellar assembly and function. The membrane plays a role in this process, as indicated by the finding that certain mutants in outer membrane proteins are nonflagellated. In E. coli, flagellar assembly appears to take place continuously throughout the cell cycle, whereas in specialized bacteria such as Caulobacter, the process has a temporal relationship with growth and the cell cycle. This difference is in keeping with the fact that E. coli has flagella located at random over its surface, whereas Caulobacter has polar flagella only.
"Examining a large number of mutants that possess partial flagellar structures under the electron microscope has revealed an elaborate choreography (Figure 17 in their book):
· The process proceeds sequentially from the proximal (= near the body) to the distal end (= farthest from body). The basal body is assembled first, then the hook, and lastly, the filament.
· The first recognizable structure is composed of the innermost rings of the basal body, incorporated into the cell membrane.
· The remainder of the basal body is then assembled and the hook is added to it, probably by extrusion through the central channel. The main protein of the filament - flagellin - plays no role in the assembly of the basal body or of the hook. Nothing is known about how the length of the hook is precisely measured, other than the fact that mutants in certain genes lead to the formation of 'superhooks.' Thus, certain proteins probably function as 'yardsticks' to determine the proper length of the hook.
· The filament is now made by extrusion of flagellin molecules through a central hollow core (Figure 18 in their book). Upon reaching the tip each molecule spontaneously condenses with its predecessors and thus elongates the filament. The phenomenon may well depend entirely on the properties of flagellin, because isolated flagellin molecules can, in the presence of a primer, self-assemble in vitro into structures indistinguishable from flagellar filaments. In vivo, the process can go on for a long time, although filament assembly slows down with increasing filament length. This slowing, plus mechanical breakage, may explain why flagella do not reach extraordinary length.
· It appears that only after the flagellum is assembled are several membrane proteins inserted near the basal body to render the structure functional. Thus, flagella cannot 'turn on' until they are fully assembled.
As far as is known, assembly of flagella occurs in a similar fashion in all bacteria. Obviously, the details of basal body formation must be somewhat different in Gram-positive and Gram-negative cells. In addition, in the spirochetes, flagellar assembly takes place entirely within the periplasm. Whereas much remains to be learned about the details of flagellar assembly, it is a particularly illuminating example of macromolecular self-assembly." Neidhardt, F. C. (1990:125-127).
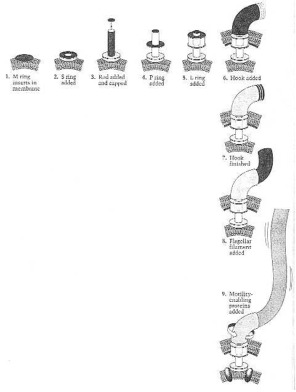
Flagellar assembly in Salmonella typhimurium. (1, 2) The M ring and the S ring of the basal body insert into the cell membrane. (3) The rod is added and the distal end of the rod is capped. (4) The P ring is added. (5) The basal body is completed by addition of the L ring. (6) The hook is made and finished (7) by addition of other proteins. (8) The flagellar filament is made. (9) Motility-enabling proteins are added to the cell membrane to finish flagellar assembly. From Neidhardt et al. (1990:126) Fig. 17.
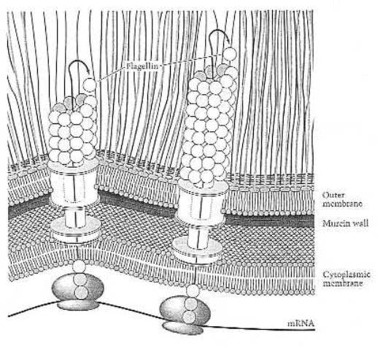
Growth of the flagellar filament by extrusion of flagellin subunits through the flagellar core. In this way the bacterial cell makes its own propeller: through automatic production, through automatic self-assembly. From Neidhardt et al. (1990:127) Fig. 18.
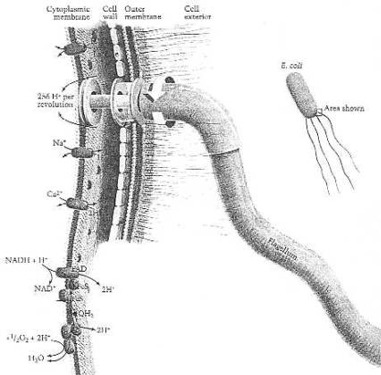
Activities of the cytoplasmic membrane that involve transport of protons. The F1 F0 ATPase is shown as it functions to synthesize ATP, the cell’s energy. From Neidhardt et al. (1990:158) Fig. 9. This is the world’s smallest rotary motor.
Prof. F. C. Neidhardt and co-workers: "The bacterial flagellum is functionally analogous to a propeller attached to a motor. Its helical filament is the propeller, its basal body with associated bearings. (The hook probably functions as a universal joint.) The motor, which is embedded in the cytoplasmatic membrane, is turned by a flow of protons driven by protonmotive force, as has been shown very directly: the flagella attached to empty cell envelopes will turn at full rate when the buffer outside the cell is adjusted to about four pH units less than that inside.
"Approximately 1,000 protons flow through the cell envelope during each revolution of the motor. This amount represents a minuscule expenditure of energy - less than 1% of the cell's energy budget is spent for motility. Although a flow of protons drives the flagellar motors of enteric bacteria and most other bacteria, other ions are sometimes used. The flagella of alkalophilic bacteria are turned by a flow of sodium ions.
"Enteric (= intestine) bacteria, being peritrochous (hairy all over), bear a number of flagella spread somewhat randomly over their surface, an arrangement that seems inconsistent with directional movement of the cell. One might imagine that such an arrangement would push the cell evenly from all directions, and that no net progress would be made in any direction. But this is not what happens. When the flagella turn in the counterclockwise direction (as viewed by looking outward from the cell) all the individual flagella coalesce into a bundle that functions as a composite propeller.
"It drives the cell evenly through the medium at a relatively rapid rate: flagellated bacteria swim at the rate of 10-20 µm/sec. Put in relative terms, this rate is equivalent to about 10 body lengths a second, a speed that would correspond to about 40 mph (64.4 km) for a human being. This condition of smooth swimming, which specialists in the field of bacterial chemotaxis call a run, does not continue indefinitely. After a brief period, the extent of which is determined by whether or not the cell is being tactically attracted, the direction of flagellar rotation reverses.
"As soon as the flagella begin to turn in the clockwise direction, the flagellar bundle flies apart and the cell tumbles without making net progress in any direction. In some still mysterious way, the turning of all flagella on a cell is coordinated: when one switches its direction of turning, they all switch. The period of tumbling is always quite brief - less than a second - but it changes the direction in which the cell swims; the next run might proceed in any direction. For taxis to occur, the tumble is as essential as the run." (1990:184, 185).
How does the bacterial cell find its food? And how does it avoid substances, which might hurt it? How does the cell smell?
Prof. F. C. Neidhardt and co-workers: "The cell measures, remembers, measures again, and compares. The cell responds to a change in the nutrient concentration, not to its absolute value, so it must constantly adapt to higher concentrations in order to be able to detect still higher ones. Indeed, adaptation is a fundamental aspect of sensory perception, our own visual sense depends on similar principles of detecting differences in light intensity at various absolute levels of intensity.
"A set of transmembrane proteins called methyl-accepting-chemotaxis proteins (MCPs) detects and measures concentrations of chemotactic compounds in the medium. As their name implies, these proteins become methylated in response to changes in concentration of attractants or repellants in their environment, and during the period of adaptation, they return gradually to their original state of methylation. A high level of methylation signals prolonged the runs, a low level increases the frequency of tumbles.
"From our discussion so far, it is apparent that MCPs must lie within an information pathway leading from environmental change to control of rotation of the flagellar motor (Figure 8 in their book). Change in the concentration of attractants in the medium determines the state of methylation of the MCPs; the state of their methylation determines how long the motor continues to turn in the counterclock direction. There are three major MCPs. - Tst, Tar, and Trg - that respond to different attractants. A particular chemo-attractant, either in a free state or bound to a periplasmatic binding protein, attaches to specific sites on the periplasma-exposed surface of its particular MCP.
"Thus, the presence of a chemo-attractant in the medium is converted into an intracellular signal registered as a methylated MCP. This signal is passed through about five intracellular proteins to a switching mechanism on the flagellar motor, causing it to prolong or diminish the time of counterclockwise rotation. How the signal is passed through the intracellular information chain and how it directs the rotation of the flagellar motor is not yet clear, but the proteins in the chain have structural similarities to proteins in two-component regulatory systems, a finding suggesting that these proteins pass information down the chain, possibly by one protein phosphorylating to the next." (1990:187, 190).
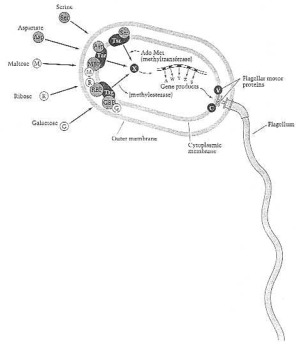
Transducer proteins and information chain leading from a chemotactic signal to control of flagellar motor. From Neidhardt et al. (1990:189) Fig. 8. The bacterial cell finds its way chemically. Its chemical sensors help it to find its food and to avoid dangerous chemicals. This sensory chemical information then controls the rotation of its flagellar motor.
What have other workers found out about the world's smallest rotary motor? How does it work?
David H. Freedman is a freelance science writer in Brookline, Massachusetts, U.S.A. He reports in Science Vol. 254, 29 November 1991 p. 1309 under the heading "Exploiting the Nanotechnology of Life":
"David Blair, a biologist at the University of Utah, has been studying the 25-nanometer-wide molecular motor that powers the propeller-like flagellum of many bacteria. Spinning at up to 18,000 revolutions per minute, the motor pushes an average-sized cell at 30,000 nanometers, or about 15 body-lengths, each second - and it is reversible too. 'It is a triumph of engineering,' gushes Blair. To tease out the motor's 'parts list,' Blair has been altering genes coding for proteins in the motor and studying the effect of each change. So far, the only part he has clearly identified is the 'fuel injector' - a proton channel that provides the motor's energy source - but he expects to be able to nail down parts corresponding to a rotor, stator, motor mount, and transmission. ... He also points out that the motor seems to be constructed of molecular rings."
The Professors T. D. Brock and M. T. Madigan report (1991:66): "The energy required for rotation of the flagellum comes from the proton motive force. Dissipation of the proton gradient creates a force, which rotates the flagellum counterclockwise and propels it through the liquid. The maximum rate of flagellar rotation is about 200 revolutions per second (12,000 rpm), and calculations of the energy consumption of the motor indicate that each flagellum draws the equivalent of about 10-15 amperes of current."
Philip Poole researches in the microbiology unit at the University of Oxford, England. He says in New Scientist 3 March 1990 p. 39 under the title "Microbes on the move": "The flagellum lies outside the cell, but it is powered by a motor that sits in the cell membrane. The word 'motor' is used deliberately: this is the world's smallest motor, at around 20 nanometres across. Unlike the more familiar motors of human technology the bacterial motor is powered by protons rather than electrons. The flagellar motor is remarkable in another respect - it causes the flagellum to rotate at around 100 hertz. Many other organisms have flagella or cilia, but these whip them back and forth, rather than rotate them as E. coli does."
Siegfried Scherer is Professor of Microbial Ecology at the Technical University of Munich, S. Germany. He states about the rotary motor of the bacterium Escherichia coli (1998:130):
Bacteria-motors of most different construction-type were found in the bacteria-world. But the motor of E. coli has been examined genetically the best. Measurements and biophysical data of the rotation-motor of E. coli, after MacNab 1996.
Measurements of the bacteria-motor
Length the cell approximately 2µm
Length the flagellum about 10 µm
Diameters the flagellum about 15 nm
Length the motor-axis about 30 nm
Equipment up to 15 motors/cell, typically: 8 motors
Technical data of the bacteria-motor
Work-tension 25-200 mVs; linear between 25-125 mV
Consumption of energy about 1200 protons/rotation 0,1% of the cell's consumption of energy
Maximum speed 100 rpm
Torque per motor about 3 10-18 Newton
Performance per motor about 10-16 Watt at 20 rpm
Maximum speed about 25 µm/sec
Corresponds to 180 km/h of a car
Production expenses 2% of the cellular biosynthesis expenditure.
The motor is driven by the energy, that is stored in the proton gradient above the cytoplasmic membrane. This proton gradient generates a surrounding, which is positively loaded opposite to the cytoplasma outside. That potential difference (= membrane potential), amounts to about 0,2 V. Pictorially spoken, the bacteria cell is a '0,2V battery'. It is able to drive the 'super nano-motor'. Scherer, S. et al. (1998:130)
Minimal demands of a primitive bacterial rotary motor
1. A first motor requires each of the following five basic elements: bacteria-flagellum, angle-piece, rotational axis, bearing and motor protein. If one of these basic elements is missing, as one can easily see, the structure, which has arisen, will not be able to function then as a motor. But it will only cost then unnecessary metabolic energy. Such a bacterium will not be able to survive in the process of selection. It will die out, and cannot be used any longer for further experiments of evolution. One has speculated that these five elements will be able to take over the function of the more than 40 proteins. But there is no molecular biological basis for this speculation. (1998:131).
2. A running motor is at a disadvantage in selection, if it cannot be steered. Therefore, also a control, that must have consisted at least of one sensor protein and a protein of signal-transmission, must have existed. On the other hand, there is no molecular biological basis to assume that these two proteins will be able to take over the function of today's approximately 8 proteins.
3. The re-buildings, which are needed for the new construction, are supposed to have taken place, by duplicating the 'pre-adapted' genes. G. Osche (1972) has formulated it aptly, when he said: A creature is not able, during the changes in the assumed process of evolution, to say, 'Because of rebuilding, temporarily closed'. Scherer, S. et al. (1998:131).
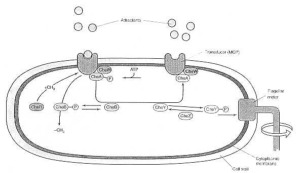
Interactions of transducers, chemotaxis (Che) proteins, and the flagellar motor in bacterial chemotaxis. The transducer (MCP) forms a complex with the sensor kinase CheA and the coupling protein CheW. This combination results in a signal-regulated autophosphorylation of CheA to CheA-P. CheA-P can then phosphorylate the response regulators CheB and CheY. Phosphorylated CheY (CheY-P) interacts directly with the flagellar motor switch. ChZ dephosphorylates CheY-P. CheR continually adds methyl groups to the transducer. CheB-P (but not CheB) removes them. The degree of methylation of the transducers controls their ability to respond to attractants and repellants and leads to adaptation. From M. T. Madigan et al. (1997:244) Fig. 7.22. From where has this biochemical and technical know-how come, which enables this tiny one-celled organism to navigate, to finds its food and to avoid dangerous chemicals?
We have looked briefly at bacteria and archaea, driven by a rotary motor, controlled by chemical navigation. We have looked at the rotary motor and its propeller in the Gram-positive and Gram-negative bacterium. It is about 20 nm across. 1 nm (nanometer) = 1 x 10-9 m (= 1/1 000 000 000 m). It propels rod-shaped bacteria, like E. coli, which is about 2 µm long and 1 µm wide (1 µm ) 1/1 000 000 m). It also moves smaller bacteria and archaea, with a diameter of 0.5 µm and less. The rotary motor, guided chemically, helps the cell, to find its food, and to avoid dangerous chemicals.
Thus, the world's smallest motor, guided chemically, serves a purpose. It is made to reach a certain goal: To keep the cell alive, and to let it become many. And many of these tiny cells serve as primary producers of food, at the beginning of the world's food-chain. This clearly disproves the doctrine of evolution, that there is neither a plan nor a goal in nature. Because thereby a Creator would be implicated.
We could compare the bacterial cell (with its spherical or rod-shaped body) with a submarine, driven by a rotary motor. This submarine is guided by chemical navigation. It helps this tiny creature to find its fuel and spare parts (the needed chemicals), and to avoid dangerous places (poisonous chemicals). This living submarine is able to repair itself. And it is able to double itself, to make two identical ones out of one, within 20 minutes to 1 hour. Compared to the bacterial and archaeal cell, even the most modern atomic powered submarine is only a clumsy thing. The ingenious design of these one-celled creatures is far beyond anything, man is able to do now.
We should also remember here: The ingenious design, found now in the bacterial and archaeal cell, has been realized already some 3.5-3.8 billion years ago, when life on earth appeared. Man has not made the bacterium and archae. He has only found them and understood them a little. No human scientist is able to make a bacterial or archaeal cell, with its rotary motor. It is far too complicated. Hence, they must have been thought out and made by someone, who has lived already long before mankind, and who knows much more about chemistry and physics, information processing, and mathematics, machine building, than any person, now living on this earth: the Creator, the God of the Bible, whose name is Jehovah. - The atheists and agnostics around the world should not ignore this fact any longer, if they want to be taken seriously. The hypothesis of Evolution (neo-Darwinism), as now commonly taught, has nothing to do with serious science. It is one of the oldest forms of false religion.
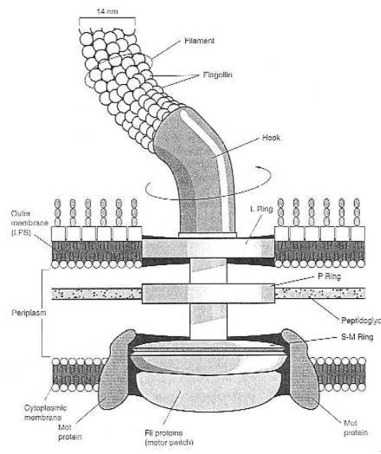
Structure of the prokaryotic flagellum and attachment to the cell wall and membrane in a gram-negative bacterium like Escherichia coli. Although cells of E. coli are peritrichously flagellated (have many propellers), for simplicity, only a single flagellum is shown. The L ring is embedded in the LPS layer, and the P ring in peptigoglycan. The S-M ring is embedded in the cytoplasmic membrane. The Mot proteins function as the flagellar motor, whereas the Fli proteins function as the motor switch. From M.T. Madigan et al. (1997:85) Fig. 3.49. Such automatic production and self-assembly of rotary motors is far beyond man’s scientific and technical ability.