
Where is the lion able to live, and where not? When will this large cat starve to death?
At the 5th of April 2002 at 19:15 o'clock in the Second Channel of the German Television Documentation, produced by the ZDF (Second Channel of German Television) and Discovery Communications, Inc., they showed a short film about the lions on the short-grass plain in the southern part of Serengeti National Park, southern Tanzania, East Africa. They showed there the so-called Goll-pride, also, how they are able to live there during the dry season. I shall quote here briefly some parts from the commentary to this film: about the so-called Goll pride. This grassy plain in the south of the Serengeti lies in the rain shade of volcanoes. During the dry season there are still a few antelopes and ostriches left. But the lion can capture them only rarely or not at all. They are too shy.
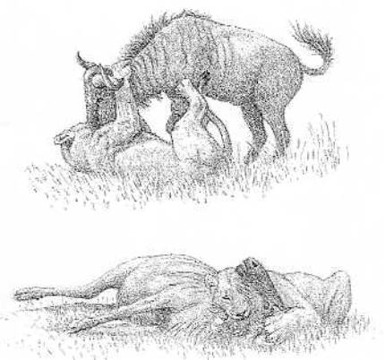
For the lions of the Goll pride, suffering from the heat, the warthogs become then the most important prey animals. These warthogs dig out now the roots and tubers of plants and devour them. Antelopes cannot dig out these roots and tubers. These warthogs come in the dry season to the last water holes, in order to drink and to wallow in them. They must wallow in there, to cool off in the tropical heat. These last water holes lie however in hollows, in the lower parts of the plain. Thus, the lion can easily crawl near to them. Now the lions of the Goll pride are living now nearly only on pork. Without the warthogs, this grassy plain in the south of the Serengeti would be practically worthless for lions. The problem is only that there are only very few pigs.
In August, during the dry season, the adult lions hardly allow their young ones to stay any longer in their pride. Even their daughters, whom they will normally allow to remain, they drive away now. Because too many lions have grown up. For the young females it becomes particularly bad in the empty, hot country. The young females try to remain within in their familiar homeland area, in which they grew up, as long as possible. But they are loosing condition now more and more. There is too little prey.
The gnus and zebras have left already some time ago the grasslands, and have wandered out of the rain-shadow of the volcanoes into the shrub country further north. Up there, there is still enough green fodder and water.
The young females have remained near the home range of their mother. The young male lions follow the gnu and zebra herds. Here they find plentiful of prey. They are living there on carrion, because they have not learned yet, to kill their own prey. That, the most experienced lionesses in his pride have done until now, in which they grew up. They must still learn that. When hunting, the lion does not hunt against the wind, that is, he does not care, from which direction the wind is blowing, despite all contrary statements.
In this film they showed then: In the tall grass of the shrub country the young male lion is crawling up meter by meter toward his prey, until he has come so close to it that he can attack: herds of gnus and zebras, which he has followed from the dry grassland in the south of the Serengeti to the north into the shrub land. An old gnu bull lost, when crossing a small body of water, in the mud the connection to the herd. The young male lion kills now this gnu bull. It is his first prey, which he has killed: The first important step into his independence. But the most important step still lies before him: He must still conquer his own home range.
His sisters in the south will probably hardly ever have a chance for this: to live on their own home range. All lions are hungering there now. The young female lions of the Goll pride are still trying to survive somehow in the no-man's land within the home range of their mother. They are only skin and bone now. Their ribs are sticking out, and their skin is shaggy. Some young animals and many old animals perish, are starving to death.
When the young males, at the beginning of the next rainy season, will come back from the north, from the damper shrub country, into the grass savanna in the drier southern part of the Serengeti, only a few residential lions, which have survived there the dry season, will welcome them,
The residential pride and the adult females, who have no home range, cannot follow the moving gnu and zebra herds. Their young ones need a well defended homeland, a home range, which has enough prey animals, also during the driest time of the year. That is, prey animals, which they can also catch.
Nomads, which have no home range of their own, follow the gnu and zebra herds. Whether the gnus and zebras will be able to survive, does not depend on the lions, but on the rain, which falls there during the year. But the long rain (the last rainy season) was not enough for this.
At the end of the dry season, of the Goll pride in the grass savanna in the southern part of the Serengeti, in their 200 km² large home range, only 6 lions have survived. Only with great difficulty were they able, to survive this dry season. The other lions have starved to death.
In another film about the lions in the south the Serengeti, in grassland, in which there are Kopjes, small rock hills, they reported this: Also a nomadic lioness, which has cubs, will lose them again. She will not be able to raise them, even if she finds there enough to eat: The male lion, controlling his home range, will kill her cubs, as soon as he finds them. Then he will take the lioness into his own pride and beget young ones with her.
In some places in the south of the Serengeti during the dry season, the grasses are nearly grazed down to the roots. In other places there is still much grass left. It is about 20-30 cm high. Why have the gnu and zebra herds then left at the beginning of the dry season this area and wandered north into the shrub country, although there was still so much grass left then? There are two reasons:
Firstly: The grassland is not able to stand it, if the hoofed animals are grazing it down there completely. The important fodder plants disappear then.
Secondly: During the dry season the grass, which still stands there, is yellowish brown and dried out. It is then like straw, with the nutritional value of old straw. Toward the end of the dry season it contains only little crude protein, about 2-2,5 percent. Elephant, buffalo and other hoofed animals will even starve to death, where the grass is 2-3 meters high, if it is dried out and if it contains too little protein. The hoofed animal can still eat it then, but will not be able to digest it anymore. It will starve to death with a full stomach. That shows us: Not only the quantity of the fodder is important, but also its quality.
A pride of lions relax in the shade of a lone tree in a grassland. The cubs are playing and feeding. From: A. Turner and M. Antón, The Big Cats (1996:152) Fig. 5.1
What is the lower limit, at which the lion will still be able to live? At which lean season prey biomass.
P. C. Viljoen, from Kruger National Park, South Africa, has studied the lions in the Chobe National Park, in Botswana, southern central Africa. He reports his finding under the title: “The effects of changes in prey availability on lion predation in a large natural ecosystem in northern Botswana”. In Symp. Zool. Soc. London (1993) No. 65:193-213. Just as in the southern Serengeti, the lion is also living in the Savuti area of the Chobe National mainly on warthogs; but there are also buffaloes.
Results of a 36-month field study on lion predation in the Savuti area of the Chobe National Park in Botswana. Buffalo and zebra were the major prey species in the rainy season (69.4%) whilst warthog, tsessbe and buffalo were most frequently killed during the dry season (7.1%). The main resident prey species was warthog.
Estimated food intake was 1.6 times higher during the rainy season than during the dry season although the observed killing rate did not change significantly. Availability of migratory prey species during the dry season appears to be the main factor limiting the size of the lion population. The presence of migratory prey during the dry season also reduced the predation effect on resident prey populations. (1993:193).
Study area: The Savuti region is situated in the western part of the 11 700 km² Chobe National Park (CNP) in northern Botswana. The main study area encompassed the Savuti Floodplain and the lower section of the Savuti Channel, a total area of approximately 300 km² between 18° 30’ and 18° 45’ S, and 24° 02’ and 24° 12’ E.
Rainfall, 550 mm average per annum, is erratic and varies considerably. Seasonal pans fill up during the rainy season (November to March) and often retain water well into the dry season (April to October.). (1993:194)
The dry-season herbivore biomass increased from 2000 kg km² to 4000 kg km² in the rainy season. However, approximately 85% of the large herbivore biomass was made up of elephant. (1993:197)
Estimated herbivore biomass in Savuti (May 1985 to February 1968); with Zebra, Warthog, Giraffe, Blue wildebeest, Tsessebe, Impala, Roan antelope, Sable antelope, Buffalo, Kudu, Eland, and Reed buck.
Rainy season /November – March): 2119.4 kg km²,
Dry season (April – October) 730.1 kg km².
Average biomass 1424.7 kg km²
Without elephant and hippopotamus. (1993:196) Table 1.
Apart from the diversity of ungulates in the region, both lion Panthera leo and spotted hyena Crocuta crocuta are numerous. Cooper (1989) estimated the hyaena population at about 393 individuals, of which approximately half were migratory. There are several leopards Panthera pardes, but both wild dog Lycaon pictus and cheetah Acinonyx jubatus are encountered only occasionally in the area. (1993:197, 198).
The meat available from each carcass is an estimate based on known dressed carcass mass (Ledger 1968; von La Chevallerie 1970). The percentage deboned meat from smaller species such as impala appears to be higher than that from larger species such as buffalo (Young & Wagener 1968). Certain body parts, for example portions of the skin and bones, are more readily eaten from smaller carcasses and an adjustment was therefore made to estimate the amount of edible meat.
|
Live mass (kg) |
Edible meat (%) |
|
< 50 |
80 |
|
50-150 |
75 |
|
151-250 |
65 |
|
> 500 |
60 |
Adapted from P. C. Viljoen (1993:199) Table 2. The larger the prey animal, the less the flesh-eater will be able to devour from it. For example: From a small antelope weighing up to 50 kg, the lion will be able to use up to 80% of its body mass. From the African buffalo the lion or other large cat will be able to use only about 60%, because it cannot eat its bones and its skin. They are too hard, too tough.
The seasonal movements of the migratory herbivore species therefore resulted in a rainy-season prey biomass that was 2.9 times higher than the dry-season biomass. The Savuti prey biomass declined from the start of the dry season in April until about June as a result of the continuous erratic movements of buffalo and zebra herds, possibly as a result of variability in rainfall. (1993:201, 202)
An average of 87.0 lions occurred in the study area (range 74-108) during the study period. A total of six permanent, resident lion prides were identified in the study area. The calculated total ranges of the six prides varied between 42 and 369 km² with an average of 217 km. The smallest range was 42 km² (Table 3).
The estimated lion biomass, based on the mass of an adult female (Bertram 1973), varied between 14.3 and 19.3 kg km². Lion biomass increased at the end of the dry season (October) whilst the lowest biomass occurred during the rainy season (January) and at the beginning of the dry season (April). Nomadic lions, the total varying between three and 24 at a time, were mainly recorded just before the start of the rainy reason and were mostly responsible for the observed changes in lion biomass. However, the resident lion biomass remained relatively stable throughout the study area (= 13.9) although the number of prides changed.
Core areas had an average size of 29 km² and 45 km² during the rainy and dry seasons respectively, with the latter on the average 1.7 times larger (range 1.2-1.8). (1993:202, 204)
The average daily meat intake was 5.6 kg in 24 hours. A higher daily intake was recorded for the rainy season (7.6 kg in 24 h) than for the dry season (4.6 kg/24 h). There was also no significant difference in the killing rate between the wet and dry seasons, 0,6 kills/24 h and 0.5 kills/24 h respectively.

Two African lions tackle a buffalo. After: A. Turner and M. Antón, The Big Cats (1996:154) Fig. 5.3.
The estimated average daily intake was 5.6 kg/24 h and is close to Schaller’s (1972) estimate of 5 kg of meat per day for a female to maintain basic metabolic requirements.
A total of 359 and 652 kills were required to meet the basic metabolic requirements of the Savuti lion population in the rainy and the dry seasons respectively. The population kill rate (kills/24 h) was 1.3 times higher in the dry than in the rainy season. It was further calculated that 1011 kills annually are necessary to sustain the lion population, of which 242 were warthog and 303 buffalo. A total of 164 zebra, 79 tsessebe and 49 wildebeest were included. P. C. Viljoen (1993:207)
Studies on lion (Schaller 1972; Bertram 1973) have demonstrated that lion populations are regulated largely by food supply and East (1984) has demonstrated that large carnivore biomass in arid/eutrophic African savannas has a positive relationship to rainfall and to prey biomass. (1993:209)
No clear evidence exists that lions or hyaenas have a limiting effect on prey in areas where prey availability is subject to seasonal changes (Kruuk 1972; Schaller 1972). With the possible exception of warthog, Svuti lions are unlikely to have a limiting effect on any of the other prey species. Cooper (1990) found that zebra constituted 80% of prey species killed by spotted hyaena in Savuti during the rainy season, but that during the dry season hyaena preyed intensively on both warthog and impala.
The resident prey population appears unlikely to provide an adequate food supply to support the same lion biomass for an extended period, particularly if the possible combined effect of hyaena predation on resident prey species is considered. An increase in the numbers of resident prey on the Serengeti plains, which followed higher rainfall, has resulted in an increase in lions, indicating that severe dry seasons might reduce the numbers of lions (Hanby & Bygott 1979).
Warthog were killed intensively during the dry season and appear to be the only species, which might be affected by predation, particularly if the hyaena predation on this species is considered as well. Migratory prey clearly provided a buffer against lion predation on resident prey during the rainy season. Furthermore, during the dry season the combined presence of tsessbe and smaller herds of buffalo and zebra, which sometimes remained in the area after the end of the rainy season, also reduced the impact of predation on warthog. P. C. Viljoen (1993:211).
Savuti region, in western part of Chobe National Park, in northern Botswana:
500 mm annual precipitation.
In rainy season 2000-4000 kg large mammal biomass.
85 % of this large herbivore biomass was made up of elephant.
Rainy season large herbivore biomass 2119.4 kg/km²
Dry season large mammal herbivore biomass 730.1 kg/km².
It appears unlikely that the dry season prey biomass of 730.1 kg/km² is able to support the present population of 87.0 lions in the Savuti region. It would not be able to feed the residential lion raising cubs. This dry season prey biomass is the nutritional bottleneck keeping the lion population at a certain level. This means, that many of the cubs and some of the older lions will starve to death during the dry season.
A lioness attacks an antelope, pulls it down, bites into its throat and chokes it to death. From: A. Turner and M. Antón (1996:168) Fig. 5.14.
The large plant-eaters and flesh-eaters on Africa’s savannas: How much biomass of large herbivores and carnivores will these tropical grasslands support? And how much must it rain there then per year? How much elephant biomass and buffalo biomass will these grassy plains be able to feed at how much rainfall per year? How much prey biomass is needed, to support which amount of large predators? What have scientists found out about this?
R. East, from New Zealand, has studied these questions. His report appeared under the title “Rainfall, soil nutrient status and biomass of large African savanna mammals” in African Journal of Ecology 1984, Volume 22, pages 254-270:
Coe et al. (1976) showed that the total biomass of large savanna herbivores was positively related to mean annual rainfall in twenty wildlife areas of southern and eastern Africa, in which rainfall varied from less than 200 to more than 1100 mm year. … In practice, herbivores are influenced by the quality as well as the quantity of available plant biomass. This is most strikingly evident in the Brachystegia/Julbernadia ‘Miombo’ woodlands which cover a vast area of the ancient elevated inland plateaus of south-central Africa. These plateaus consist largely of pre-Cambrian rocks and weather into highly leached soils with very low supplies of nutrients available for plant growth; the resultant poor-quality vegetation supports a low herbivore biomass (Brown, 1965). Miombo woodlands occur mainly in areas with mean annual rainfall of 850-1400 mm, but they support a much lower biomass of large herbivores than would be predicted from the biomass/rainfall relationship derived by Coe et al (1976).
Bell (1982) showed that the herbivore biomass/rainfall relationship is modified by geomorphology through its influence on the availability of soil nutrients. He suggested that there is a specific biomass/rainfall relationship for each particular geomorphological situation and hence class of soil nutrient availability. Whereas large herbivore biomass increases with rainfall on soils of volcanic origin (high soil nutrient status) and rift-valley sediments (medium soil nutrient status) as mean annual rainfall increases from less than 200 to more than 1000 mm, in basement situations (low soil nutrient status), as on soils derived from ancient granitic continental shields, biomass increases with rainfall up to a mean annual rainfall of about 700 mm and then appears to decline as rainfall increases further (Bell 1982). R. East (1984:246)
The biomass of some species tends to be lower in moist/dystrophic savannas than in arid/eutrophic [fertile] savannas over the same rainfall range, 700 to more than 1000 mm per annum, e.g. elephant and buffalo. This trend is not apparent for other species, such as the hartebeest (Alcelaphus buselaphus). Herbivores can be divided into two arbitrary groups, ‘arid’ and ‘moist’ savanna species. According to whether estimates of their biomass on soils of low nutrient status peak at annual rainfalls < 820 mm or >1000 mm respectively.
The biomass of arid savanna herbivores on soils of low nutrient status increases with rainfall to reach a peak at an annual rainfall of less than 820 mm (less than 600 mm in most cases) and tends to decline as rainfall decreases further –e.g. elephant, zebra, buffalo, giraffe, black rhinoceros and eland. As a result, the biomasses of these species on low nutrient status soils show no significant relationships to rainfall over the full range of rainfall, … Some tend to be particularly abundant on the medium nutrient status soils of rift valley habitats –e.g. elephant, black rhinoceros and impala (Aepyceros melampus).
Arid savanna species are generally widespread – i.e. have an occurrence of at least 50% in arid/eutrophic savanna reserves where annual rainfall exceeds 350 mm. With the exception of elephant, hippopotamus, buffalo and eland, the occurrence of these species in moist/dystrophic savannas declines markedly as rainfall increases above 1000 mm –e.g. giraffe, wildebeest (Connochaetes taurinus), greater kudu and impala. Oryx, springbok, Grant’s and Thomson’s gazelles are largely or entirely absent from moist/dystrophic savannas. The topi and conspecific tsessebe are patchily distributed. R. East (1984:250).
![]()
A pride of female lions attack a zebra. From: A. Turner and M. Antón, The Big Cats (1996:153) Fig. 5.2.
Small herbivores (unit weight 15-60 kg) range in size from Thomson’s gazelle and oribi to warthog, impala and kob. Medium-sized herbivores (unit weight 61-450 kg) range from larger antelopes such as topi, hartebeest, waterbuck and roan up to buffalo. Very large herbivores (unit weight >750 kg) – viz. elephant, rhinoceroses, giraffe and hippopotamus –are not included in Table 4 because they are not often preyed upon by carnivores.
Although few data are available, total large carnivore biomass is positively correlated with rainfall in arid/eutrophic savannas and with total prey biomass in all areas. The biomass of each size class of prey and the biomasses of most individual carnivore species (Table 4) are also positively correlated with rainfall in arid/eutrophic savannas. Carnivore species differ in the size range of prey normally taken. In his review of predation studies Eltringham (1979) showed that the lion (Panthera leo) most commonly preys on larger ungulates up to the size of buffalo, the spotted hyaena (Crocuta crocuta) takes a similar size of prey at last in areas where it hunts in packs, and the leopard (Panthera pardus), cheetah (Acinonyx jabatus) and wild dog (Lycaon pictus) concentrates on smaller prey up to the size of an adult impala (45 kg). R. East (1984:257, 258)
Biomass/rainfall relationships for individual herbivore species.
It is clear from Fig. 1 that the positive correlation between total large herbivore biomass and rainfall observed by Coe et al. (1976) in arid/eutrophic savannas also applies in general to individual herbivore species. …
The population of a herbivore in a particular area may fluctuate in the long term in response to changes in the structure and biomass of both the plant and animal communities with variations in the balance between water and soil nutrient availability (Bell, 1982). Population interactions between such factors as weather, vegetation, fire and other members of the herbivore community and in some cases predators. (1984:259, 261).
Moist/dystrophic [infertile] savannas: The low biomass of herbivore communities in moist/dystrophic savannas reflects the limitations imposed by vegetation quality rather than quantity (Bell, 1982). (1984:262)
Body size has an important influence on the ecology of savanna herbivores. Smaller species require a higher quality diet because of their higher metabolic rate per unit body tissue (Bell, 1971). (1984:263)
Carnivore biomass: Resources ultimately limit the tertiary trophic level. Studies of individual large carnivore species such as lion and spotted hyaena have shown that their populations are regulated by the food supply, in the case of the lion through the intermediary of social behaviour. The predator’s food supply may not be related to prey biomass in a simple way because of the influence on hunting success of such factors as the migratory habits of the prey and the amount and nature of the plant cover. R. East (1984:265)
|
Rainfall mm/year |
Total biomass kg/km² |
Rainfall mm/year |
Total biomass kg/km² |
|
100 |
78 |
600 |
8250 |
|
150 |
200 |
650 |
8400 |
|
200 |
370 |
700 |
8500 |
|
250 |
500 |
750 |
8700 |
|
300 |
2000 |
800 |
8800 |
|
350 |
3500 |
850 |
9000 |
|
400 |
5000 |
900 |
9150 |
|
450 |
7600 |
950 |
9300 |
|
500 |
7600 |
1000 |
9455 |
|
550 |
8100 |
1500 |
9620 |
Adapted from R. East (1984) Fig. 3a. Total biomass of African arid savanna herbivores and mean annual rainfall. Soil nutrient status: high. In: “Rainfall, soil nutrient status and biomass of large African savanna mammals” in African Journal of Ecology 22:245-270 (1984)
Biomass of African Buffalo and annual rainfall
|
Rainfall mm/year |
Buffalo biomass kg/km² |
Rainfall mm/year |
Buffalo biomass kg/km² |
|
50 |
50 |
700 |
910 |
|
100 |
100 |
750 |
962 |
|
150 |
130 |
800 |
2400 |
|
200 |
200 |
850 |
3900 |
|
250 |
219 |
900 |
5300 |
|
300 |
400 |
950 |
6850 |
|
350 |
500 |
1000 |
8342 |
|
400 |
600 |
1050 |
8440 |
|
450 |
710 |
1100 |
8550 |
|
500 |
820 |
1150 |
8700 |
|
550 |
820 |
1200 |
8800 |
|
600 |
880 |
1250 |
8950 |
|
650 |
900 |
1300 |
9100 |
Adapted from R. East (1984) Fig. 1. Biomass of African buffalo and mean annual rainfall. In: “Rainfall, soil nutrient status and biomass of large African savanna mammals” in African Journal of Ecology 22:245-270 (1984)
For example: at 1300 mm rainfall per year, there will be up to 9100 kg/km² buffalo biomass on the African savanna, but 9500 kg/km² elephant biomass (see, please, next table below). That is 400 kg/km² more. Why? Because the elephant is much larger than the buffalo and does have, therefore, a lower metabolic rate. At a lower metabolic rate the elephant needs less food per kilogram of metabolic bodyweight per day, than the buffalo. Thus, the same amount of rainfall and plant production is able to support a little more elephant biomass than buffalo biomass.
Biomass of African Elephant and annual rainfall
|
Rainfall mm/year |
Elephant biomass kg/km² |
Rainfall mm year |
Elephant biomass kg/km² |
|
100 |
50 |
750 |
6897 |
|
150 |
80 |
800 |
7300 |
|
200 |
95 |
850 |
7700 |
|
250 |
110 |
900 |
8100 |
|
300 |
260 |
950 |
8500 |
|
350 |
400 |
1000 |
8925 |
|
400 |
600 |
1050 |
9000 |
|
450 |
750 |
1100 |
9100 |
|
500 |
919 |
1150 |
9200 |
|
550 |
2100 |
1200 |
9300 |
|
600 |
3300 |
1250 |
9391 |
|
650 |
4500 |
1300 |
9500 |
|
700 |
5700 |
1350 |
9600 |
Adapted from R. East (1984) Fig. 1. Biomass of African elephant and mean annual rainfall. In: “Rainfall, soil nutrient status and biomass of large African savanna mammals” in African Journal of Ecology 22:245-270 (1984).
For example: at 1300 mm rainfall per year, there will be up to 9500 kg/km² elephant biomass on the African savanna, but only 9100 kg/km² buffalo biomass (see, please, other table above). Why? Because the elephant is much larger than the buffalo and does have, therefore, a lower metabolic rate. At a lower metabolic rate the elephant needs less food per kilogram of metabolic bodyweight per day than the buffalo. Thus, the same amount of rainfall and plant production will be able to support a little more elephant biomass than buffalo biomass.
![]()
African lioness with her small cub, resting. The coat of her cub is still spotted. From: A. Turner and M. Antón, The Big Cats (1996:94) Fig. 4.1.
Total biomass of large African carnivores and mean annual rainfall
|
Mean annual rainfall mm |
Total large carnivore biomass kg/km² |
Mean annual rainfall mm |
Total large carnivore biomass kg/km² |
|
50 |
1.2 |
550 |
14.0 |
|
100 |
1.5 |
600 |
15.0 |
|
150 |
2.5 |
650 |
22.0 |
|
200 |
3.1 |
700 |
22.5 |
|
250 |
4.0 |
750 |
25.0 |
|
300 |
5.1 |
800 |
20.5 |
|
350 |
5.5 |
850 |
22.5 |
|
400 |
9.0 |
900 |
34.0 |
|
450 |
10.5 |
950 |
37.0 |
|
500 |
12.0 |
1000 |
40.0 |
Adapted from R. East (1984) Fig. 4a. Total biomass of large African carnivores (15-450 kg body weight) and mean annual rainfall. In: “Rainfall, soil nutrient status and biomass of large African savanna mammals” in African Journal of Ecology 22:245-270 (1984)
Total biomass of large African herbivores and large carnivores
|
Prey biomass kg/km² |
Large carnivore biomass kg/km² |
Prey biomass kg/km² |
Large carnivore biomass kg/km² |
|
50 |
0.30 |
2050 |
12.80 |
|
100 |
0.61 |
2100 |
13.10 |
|
150 |
0.90 |
2150 |
13.40 |
|
200 |
1.25 |
2200 |
13.70 |
|
250 |
1.60 |
2250 |
14.00 |
|
300 |
1.90 |
2300 |
14.40 |
|
350 |
2.20 |
2350 |
14.70 |
|
400 |
2.50 |
2400 |
15.00 |
|
450 |
2.80 |
2450 |
15.30 |
|
500 |
3.10 |
2500 |
15.60 |
|
550 |
3.45 |
2550 |
15.90 |
|
600 |
3.75 |
2600 |
16.20 |
|
650 |
4.10 |
2650 |
16.50 |
|
700 |
4.40 |
2700 |
16.80 |
|
750 |
4.70 |
2750 |
17.10 |
|
800 |
5.00 |
2800 |
17.40 |
|
850 |
5.30 |
2900 |
18.00 |
|
900 |
5.60 |
3000 |
18.60 |
|
950 |
5.90 |
3500 |
22.00 |
|
1000 |
6.25 |
4000 |
25.00 |
|
1050 |
6.50 |
4500 |
28.00 |
|
1100 |
6.90 |
5000 |
31.00 |
|
1150 |
7.20 |
5500 |
34.00 |
|
1200 |
7.50 |
6000 |
37.00 |
|
1250 |
7.80 |
6500 |
40.50 |
|
1300 |
8.10 |
7000 |
43.60 |
|
1350 |
8.10 |
7500 |
47.00 |
|
1400 |
8.70 |
8000 |
50.00 |
|
1450 |
9.00 |
8500 |
53.00 |
|
1500 |
9.40 |
9000 |
57.00 |
|
1550 |
9.70 |
9500 |
60.50 |
|
1600 |
10.00 |
10,000 |
64.50 |
|
1650 |
10.30 |
10,500 |
70.00 |
|
1700 |
10.60 |
11,000 |
76.00 |
|
1750 |
10.90 |
11,500 |
82.00 |
|
1800 |
11.30 |
12,000 |
88.00 |
|
1850 |
11.50 |
12,500 |
94.00 |
|
1900 |
12.00 |
|
|
|
1950 |
12.20 |
|
|
|
2000 |
12.50 |
|
|
Adapted from R. East (1984) Fig. 4b p. 259. Total biomass of large African herbivores and of large carnivores weighing 15-450 kg. This shows us, how large the biomass of the prey animals must be, so that it will be able to feed a certain amount of biomass of large flesh eaters. That is, how many large predators like lions, hyenas, and leopards (in kg/km²) will be able to live then on Africa’s savanna.