Chapter 1: The Tiger of the Mammoth Fauna
The German paleontologist Ralf-Dietrich Kahlke says about the tiger of the upper Pleistocene in his book, Die Entstehungs-, Entwicklungs- und Verbreitungsgeschichte des oberpleistozänen Mammuthus-Coelodonta-Faunenkomplexes in Eurasien (Großsäuger) (1994):
“During the upper Pleistocene, the range of the northern populations of P. tigris reached from N- and NE-China to the Salawu area. … Remains have also been found in the southern Primorskiy Krai of the Russian Federation, and from N- and S-Korea. … The possibility that the tiger has spread out during the upper Pleistocene even into eastern Beringia (= Alaska, Yukon Territory) has been considered by Herrington (1987: 55). Generally, it may be said, that a relatively large northern form of the tiger (P. tigris ssp.) as an element of the Far Eastern fauna, has lived in biotopes, covered more or less with forests, as a part of the upper Pleistocene Mammuthus-Coelodonta faunal complex.” (1994:24).
“The northernmost tiger population from the upper Pleistocene, reported so far, has lived mainly in taiga-like mixed-forest-biotopes of N- and NE-China, and the Primorski Krai, in the mountain chains of Heilong-jang. It also went into the mountain-forests. Towards the NW, the more open lion-biotopes began. For the northern mainland tiger, at least during the Last Glacial, a certain cold-resistance must be assumed. The greatest depth of the snow-cover would have been – like today – around 0,3 m. Deeper layers of snow do affect the density of potential prey animals. That is, why also the tiger avoids these areas.” Kahlke, R.R. (1994:76).
Sandra J. Herrington, Museum of Natural History, University of Kansas, Overland Park, Kansas reports about her findings under the title, “Subspecies and the Conservation of Panthera tigris: Preserving Genetic Heterogeneity”. In Ronald L. Tilson and Ulysses S. Seal (eds.) Tigers of the World 1987 Park Ridge, New Jersey, U.S.A. She states about the fossil history:
“Fossils definitely assigned to P. tigris are from the lower to upper Pleistocene. These are from central Asia (Brandt 1871, Tscherski 1892), eastern and northern China (Hooijer 1947, Loukashkin 1938), northern Siberia (Tscherski 1892), Sumatra (Brongersma 1937) and Java (Brongersma 1935) and Japan, Hemmer pers. comm.). Very late Pleistocene to early Holocene fossils are recorded from the Caucasus (Vereschagin 1959) and India (Lydekker 1886). The fossils from northern Siberia ranged up to 70° latitude. These species are of somewhat uncertain affinity, and have also been allied with cave lions (P. spelaea; Heptner and Sludskii 1972).
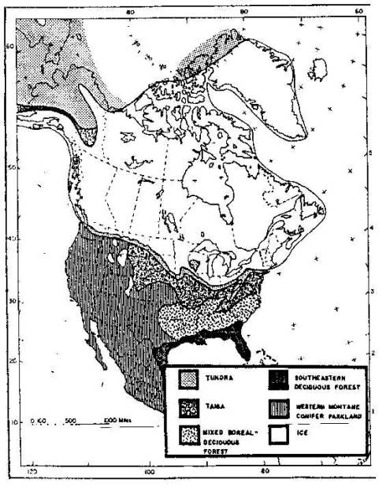
“The possible presence of tigers this far north, at a time when the Beringian subcontinent linked north-eastern Asia and north-western North America, raises the questions of why tigers did not cross the Bering land bridge, when so many other mammals did. Tigers appear to shun open country (Seidensticker 1976), and the Bering land bridge appears to have been primarily steppe tundra habitat (Fig. 2) However, there is evidence that a variety of habitats were to be found in Beringia, including wooded regions (Hopkins 1982).
“There were large Panthera in North America during the Pleistocene (Fig. 3), and these were assigned to the Fossil lion species (P. atrox; now considered by some to be a sub species of P. leo or consubspecific with cave lions; e.g., Kurten 1985). However, it is not always easy to distinguish between modern lions and tigers, based only on skeletal morphology, so the possibility remained that tigers were among the Northern American Ice Age Panthera in the past (e.g. Merriam and Stock 1932). These fossils were compared with lions and tigers to determine their affinities. However, the possibility that both lions and tigers were inadvertently included in P. atrox seems never to have been considered.
“In a recently completed study of these fossils, I developed a set of morphometric characters that distinguished between lions and tigers with 100% accuracy in discriminant function analyses. In addition, there were several qualitative morphological characters that could distinguish P. leo and P. tigris with a high degree of accuracy. I than compared modern lions and tigers with the fossil North American material. All fossil material from the area of the contiguous United States, an area south of the continental ice sheet during the Wisconsin glaciation, represented lions.
“However, the material from Beringia included both lions and tigers. It appears that lions invaded eastern Beringia during the Illinoian glaciation, and arrived in the area of the contiguous United States during the Sangamonian interglacial. Tigers, and a different population of lions, penetrated as far as eastern Beringia during the Wisconsin glaciation in a separate dispersal event. These results suggest that other large fossil Panthera may have been misidentified, and additional evaluation may further increase the range of fossil tigers.” Herrington, S. J. (1987:53-55)
“Unlike the fossil lions, the fossil tigers of Beringia were not larger than their modern counterparts. Fossil tigers from southern China from the collection of the American Museum of Natural History (Hooijer 1947) appear to be larger than modern P. tigris but only slightly so.” Herrington, S. J. 1987:56).
North America during the height of the Last Ice Age. Map from S. J. Herrington. In: Ronald L. Tilson and Ulysses S. Seal (eds.) Tigers of the World (1987:54) Fig. 2. South of the North American Ice Sheet, they have found only the remains of the lion (Panthera atrox), no remains of the tiger. Northeast of the Continental Ice Sheet, in the Yukon Territory, Alaska, Northeast Siberia and northern Central Siberia, they have found the remains of the lion and the tiger. These large cats have lived in the Far North in a temperate climate, when the woolly mammoth was grazing up there.