
Chapter 6: Short-faced Bear 2
Palaeontology Program, Government of the Yukon. Occasional Papers in Earth Sciences No. 7. Locomotor adaptations and ecomorphology of short-faced bears (Arctodus simus) in Eastern Beringia
By Paul E. Matheus, The Alaska Quaternary Center, Department of Geology and Geophysics and The Institute of Arctic Biology, University of Alaska Fairbanks. Yukon Palaeontology Program, Department of Tourism and Culture, Elaine Taylor, Minister, 2003.
Abstract
“The giant short-faced bear (Arctodus simus) was a widespread Tremarctine (New World) bear indigenous to North America until its extinction around 11,500 BP. Arctodus inhabited Pleistocene ice-free refugia in Eastern Beringia (the northwestern limit of its range) until at least 20,000 BP. Arctodus was the largest bear and perhaps the largest species of terrestrial Carnivora that ever lived, yet it was characterized by a gracile post-cranial morphology and it was relatively long-legged.
Previously (Matheus 1994, 1995) I extracted fossil bone collagen from east Beringian short-faced bears, brown bears, and other carnivores for stable isotope analysis (δ 13C and 15N) in order to test competing dietary models and the competition hypothesis. Results of that study showed that Arctodus was highly carnivorous, that its diet was completely terrestrial (i.e., no salmon), and that if fed on herbivores, which consumed C3 vegetation. The herbivore/omnivore foraging model was thus rejected. (2003:VII)
Short-faced bears were extremely large, even for bears (Fig. 1 and 2), and various attempts have been made to estimate the body weight of individual specimens. Most of these estimates were calculated using allometric equations relating long bone cross-sectional area to body mass, while some are best guesses based on comparisons to other bears. Generally, these and Corner (1986) even suggested that large males may have topped 1000 kg. Kurtén (1967a) showed that sexual dimorphism was pronounced, as it is in other bears, on the order of 15 – 25 %; this would place females to be around 450 – 600 kg. If Voorhies and Corner’s extreme estimate (1000 kg for males) is correct, then some females may have even reached 750 kg. Considering Arctodus’ gracile build, and the mass of modern bears, I think that Voorhies and Corner’s estimates are too extreme. … Furthermore, Voorhies and Corner do not explain how they derived their estimate of 1000 kg for large males in their 1986 paper, but based on their 1982 article it seems that they arrived at this value simply by their impression with how big Arctodus long bones are compared to those of modern black bears (Ursus americanus)
Even if one accepts a conservative estimate of 500 kg and 600 kg for an average female and male short-faced bear, respectively, this means that an average individual was nearly as massive as a small domestic horse and two to three times the mass of a modern, non-coastal grizzly bear (Ursus arctos horribilis). Figure 1b compares the skeletal size of Arctodus (a) to the morphs of male brown bear: the largest known modern, male, brown bear (which could also represent the largest known polar bear) (b); a very large modern, male, brown bear from coastal Alaska or Kamchatka (also the size of a typical large brown bear from interior Alaska during the Pleistocene) (c), and: a large, modern, male brown bear typical of interior Alaska, the contiguous 48 United States, Europe, and interior Russia (d). Figure 2 compares the size of Arctodus to other carnivores that are discussed.
Comparisons to other bears, however, can be misleading because Arctodus was not built like more familiar modern bears. In contrast to the stereotypical impression of a bear’s morphology, Arctodus had a very light build, with a laterally compressed but deep thoracic cavity, and limb bones that were very long, gracile, and lacked much of the characteristic bowing and toe-in posture of ursine bears.” Paul E. Matheus Quaternary Research 44 (1995:1, 2)
From: Paul E. Matheus Quaternary Research 44 (2003) Figure 1. A) Reconstruction of the short-faced bear’s skeleton. B) Size comparison between short-faced bears and various brown bears: a = large male short-faced bear, b = largest size attained by coastal brown bears today (could also represent the largest polar bear), c = large male brown bear typical for eastern Beringia during the Pleistocene (also approximate size of a large male modern coastal brown bear), d = large male grizzly from modern interior Alaska (units in meters) (original illustrations by author).
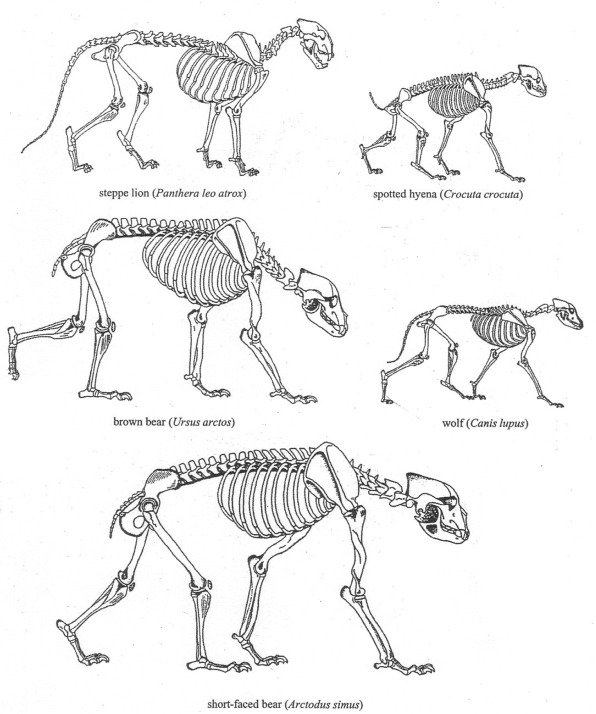
From: Paul E. Matheus Quaternary Research 44 (2003) Figure 2. Body size and skeletal shape of short-faced bears and other carnivores discussed in text. All body sizes adjusted to depict adult males in Pleistocene east Beringia, except hyena, which did not occur in Beringia. The brown bear (Ursus arctos) has lived in eastern Beringia (Alaska and Yukon Territory), when the woolly mammoth was grazing up there. This brown bear has lived then in central Alaska and the Yukon Territory. It was just as large and as heavy as the large brown bear living now at the southern coast of Alaska.

From: Paul E. Matheus Quaternary Research 44 (2003) Figure 21. A large male short-faced bear standing upright on his hind legs.
How much food did the short-faced bear need per day and per year, the adult male and female?
A large 800 kg male short-faced bear would need 17.6 kg (17561 g) of food eaten every day, or 6424 kg per year. A 600 kg female would require 5271 kg/year and the population average (assuming 700 kg) would be 5853 kg/year. (2003:86)
The following values for herbivore body mass will be used:
Mammoth 3800 kg
Bison 650 kg
Caballine horse 75 kg
This is the mean adult body mass averaged for males and females.
Woolly mammoth male 4900 kg, and female 2700 kg. (2003:87)
One short-faced bear needs then per year:
2.0 woolly mammoth, or
12.0 bison, or
44.6 caballine horses.
This includes edible meat and non-edible bones and food in intestines.” (2003:88)
“Earlier, I estimated that the average short-faced bear (700 kg) needed to consume 5853 kg of flesh per year, equal to about 16 kg per day. That is 100 kg of edible carrion every 6.25 days. If its home range was 1000 km²/year, its habitat would have to produce then a carcass biomass of at least 5.87 kg flesh/km²/year. (2003:91)
In addition to short-faced bears, fossils of large carnivores from the region [Alaska and Yukon Territory] include wolf (Canis lupus), lion (Panthera leo atrox), brown bear (Ursus arctos), wolverine (Gulo gulo), scimitar (sabertoothed) cat (Homotherium serum), and dhole (Cuon alpinus) (listed in order of descending abundance).
Patterns of fossil abundances also led me to conclude that wolves and lions were the only significant, long-term, predators that occupied eastern Beringia during the late Pleistocene. The other carnivores either are not significant predators (brown bear, wolverine), or appear to have a limited (or at least unknown) chronological range in Beringia (scimitar cat, dhole).
Brown Bear: Brown bears migrated to eastern Beringia from Asia probably in the early Wisconsinan Glaciation. But they should not be considered significant predators in the system. Stable isotope rations (C, N) in fossil brown bears from eastern Beringia indicate that they ate mostly vegetation, but also varying amounts of terrestrial meat (Matheus 1994, 1995). Today, no population of brown bears relies primarily on carnivory, even though predation and scavenging can be locally or seasonally important. Predation by brown bears on ungulates in Alaska, for instance, occurs mostly during the brief period of calving seasons when caribou, moose and occasionally sheep are taken. At these times, brown bear predation can be intense, and the biomass harvested is relatively small. But being adept scavengers, brown bears will compete with other carnivores for existing carcasses throughout the year, and brown bears will cache and defend carcasses. (2003:92, 93)
Lion: Guthrie (1990a) stated that lions probably were Beringia’s most prevalent carnivore (based on fossil abundance). However, based on the number of fossils in the Alaska collection of the American Museum of Natural History it seems that wolves may have been just as common, or more so. Lions are highly predatory but they readily turn to scavenging when it is beneficial and available (Kruuk 1972, Schaller 1972).
Schaller (1972) found that Lions in the Seronera era of the Serengeti consume approximate 2500 kg of prey per lion annually, yielding a daily individual consumption rate of 6.8 kg. The mean body mass of five female and 14 lions weighed by Schaller was 166 kg. For this weight, the expected daily food consumption predicted by Harestad and Bunnel’s equation (Equation 22) is 6.0 kg per day.” Paul E. Matheus Quaternary Research 44 (2003:94)
|
Ecosystem (type of biomass reported) |
Large mammal biomass (kg/km²) |
Source |
|
Entire Serengeti Plain (large herbivore prey) |
Annual = 4222 Zonal range = 998 – 7234 |
Schaller (1972) |
|
Woodlands of western Serengeti (large herbivore prey) |
3110 |
Bell 1967 (in Schaller 1972) |
|
Savannah of Kruger Park (large herbivore prey minus elephant) |
7785 |
Watson and Turner 1965 in (Schaller 1972) |
|
Grassland and savannah of Ngorongoro Crater (large herbivore prey) |
16,200 |
Kruuk 1972 |
|
Ruwenzori Park, Uganda plains tropical forest (large herbivores, also many elephants and hippos) |
20,000 |
Eltringham 1974 (in Redmann 1982) |
|
Boreal forest of Alberta (forest steppe) (large herbivores) |
5700 |
Telfer and Scotter 1975 (in Redmann 1982) |
|
Parkland of Alberta (large herbivores) |
2530 |
Flok 1970 (in Redmann 1982) |
|
Dry tundra of St. Elias region, Yukon (large mammals) |
89 (actual estimated) 175 (potential estimated) |
Geist 1978 (in Redmann 1982) |
|
Tundra of Southampton Island, Northwest Territories (caribou), northern part of Hudson Bay |
36 |
Parker 1975 (in Redmann 1982) |
|
Tundra of Alaskan arctic slope (caribou, Western Arctic Herd) |
91 |
Ballard et al. 1990 |
Adapted from: Large mammal standing biomass of modern ecosystems. Paul E. Matheus Quaternary Research 44 (2003) Table 13. Comment: This shows us, how large the biomass of hoofed animals is, where the residential lion, raising cubs, is able to live now in Africa. Such a large prey biomass of hoofed animals is able to live now in North America only in the forest-steppe or aspen-parkland. That is, where the steppe (prairie) changes into the boreal forest (taiga).
The dry tundra of the St. Elias region, southwestern Yukon, in the rain-shadow of the Coast Range, is supposed to be very similar to the mammoth steppe of eastern Beringia (Alaska and Yukon), when the mammoth was grazing up there. It has now a biomass of hoofed animals of 89 kg/km², and is able to support up to 175 kg/km². But we must realize here: This biomass of hoofed animals consists of Dall sheep. And these mountain sheep are able to live only at a few isolated spots, where there are steep cliffs and enough pasture ground. So, actually, the biomass of hoofed in the whole area is much lower.
The tundra of the Alaskan arctic slope, at the Alaska’s arctic coast, has a biomass of hoofed animals of 91 kg/km². It consists of caribou, of the Western Arctic Herd. But this caribou herd will live up there only in summer, when calving. In autumn these caribou will wander further south into the boreal forest, into the taiga. Then the biomass of hoofed animals on the tundra of the Alaskan North Slope will be near zero. So this will not help a lion. The lion was only able to survive in Alaska and the Yukon, when the mammoth was grazing up there, when there is also enough food during the leanest time of the year.
The residential lion, raising cubs, needs a prey biomass of hoofed animals of at least 1000 to 5000 kg/km². When the lean season prey biomass is less, the cubs will starve to death. Also the Siberian tiger shows us: Nomadic tigers will move up much further north in Siberia, than the residential tiger, raising cubs. There they are able to live up there for many years. But the prey biomass is so low, that the tiger does not reproduce, it does not get any cubs. Neither the lion nor the short-faced bear were able to live in Alaska and the Yukon as nomads. They are only able to survive, where they live as residents on a home range, raising their cubs. And that is only possible, where the lean season prey biomass is large enough. Any surplus of ungulates during the rest of the year is irrelevant.
The lion, mammoth, and short-faced bear are supposed to have also lived in Alaska and the Yukon during the height of the Last Ice Age, in “refugia”, not covered by ice. That is, when most of North America was covered by large inland ice-sheets. In the western Yukon Territory and Central Alaska, the animals of the mammoth fauna are supposed to have lived on a mammoth steppe or tundra-steppe. The tundra of Southampton Island, Northwest Territories, is similar to this mammoth-steppe or tundra-steppe, when the ice-sheets covered most of North America. It lies at the northern part of Hudson Bay, at about the latitude (height) of Dawson City and Fairbanks. The biomass of hoofed animals (caribou) is there now only 36 kg/km². Neither the lion nor the short-faced bear would be able to live now up there. They would starve to death.
Redmann’s (1982) Model
“Redmann’s model estimates the biomass density of large herbivores in a northern grassland using the following assumption: primary productivity of edible above ground biomass is set at 150 kg/km²; this is the amount of forage (‘herbage’ in Redmann) available to herbivores and is a conservative approximation based on productivity rates measured in modern cold-temperate grasslands.” Paul E. Matheus Quaternary Research 44 (2003:97, 98)
“Table 14 shows the resulting population densities calculated for this theoretical system assuming a total standing large herbivore biomass of 5100 kg/km²/year. Results show that this system would simultaneously support population densities of 0.402 mammoth/km², 3.138 bison/km², 5.246 caballines/km², and still give another 612 kg/km² divided between other minor species (e.g., caribou, muskox, saiga, elk, camel, etc.). this latter amount would support 2.5 individuals/km² of a species weighing 250 kg, for instance.
|
Species (mass in kg) |
% of biomass |
Resultant biomass density (kg/km²) |
Resultant population density (animals/km²) |
Total population in eastern Beringia (= pop. Density x 2,000,000 km² |
|
Mammoth (3800) |
30 |
1527 |
0.402 |
804,000 |
|
Bison (650) |
40 |
2040 |
3.138 |
6,276,000 |
|
Horse (175) |
18 |
918 |
5.246 |
10,491,429 |
|
Others (= 250) |
12 |
612 |
2.448 |
4,896,000 |
From: Paul E. Matheus Quaternary Research 44 (2003:100) Table 14. Estimated population densities and total numbers of dominant large herbivores in eastern Beringia (Alaska and Yukon) according to a modified biomass estimate using Redmann’s (1982) model. Total biomass predicted by the modified model is 5100 kg/km², which was divided amongst each species according to relative biomass estimates modified from Guthrie (1968).
This means: When the mammoth was grazing in Alaska and the Yukon, there had to be a biomass of hoofed animals of 5097 kg/km². That is correct. I do agree with this. Because only at this amount of hoofed animals could the residential lion, raising cubs, have lived. But this biomass of hoofed animals could not have lived up there in an arctic climate. Because of at least two reasons: 1) Aboveground plant production would have been too low. 2) During the long arctic winter the aboveground vegetation would have contained too little digestible crude protein.
The mammoth would have starved then in winter due to lack of energy and protein. This fine model will only work much further south, in the meadow-steppe (tall grassland) and forest-steppe (aspen parkland) of Alberta, where the steppe (grasslands) changes over into the boreal forest (taiga). That is about 1200 km further south. That is, when this animal fauna does not contain the mammoth.
Paul E. Matheus concludes that the biomass of hoofed animals in eastern Beringia, when the mammoth was grazing up there, should be much lower. He believes that this mammoth steppe or tundra steppe had a biomass of large hoofed animals of only 1000 kg/km². For details see his Table 16 in 2003:103, 104. Also that will not work in an arctic climate. That is completely unrealistic.