Pourquoi sont des cellules vivantes? Pourquoi est-ce qu'ils sont capables
de vivre? Pourquoi est-ce que la cellule est capable, faire ses propres jeux de
construction? Comment a compliqué est-ce que ses réactions chimiques sont? Si
vous, cher lecteur, n'en comprenez pas quelques-uns des termes techniques (ou
même la plupart d'eux), ne vous inquiétez pas vers lui. Dans ce chemin, je veux
vous montrer juste, cela qui la cellule minuscule que tout savent vers
biochimie, et ce que c'est capable de faire. Alors demandez-vous, s'il vous
plaît: Pourquoi est-ce que ce petit organisme uniloculaire sait tout ce vers
biologie moléculaire et biochimie, ce qu'un professeur de biologie moléculaire
et biochimie a dû apprendre en premier? Beaucoup des détails techniques dans la
cellule vivante sont compliqués ainsi, que les experts eux-mêmes ont été
capables, en comprendre maintenant seulement quelques-uns. Quelques-uns ils ont
été capables, comprendre seulement un peu, et quelques-uns, pas à tout. Au
moins pas jusqu'à maintenant.
Bruce Alberts est professeur américain de Biologie. Il est Président de l'Académie
Nationale de Sciences (NAS) d'Amérique. Lui et ses collègues croient que la vie
a évolué par elle-même sur terre. Ils égalisent réclamez dans leur manuel
scolaire The Cell (1989:41): Il n'y a
aucune pièce dans la vue contemporaine de vie... pour n'importe quoi autrement
à l'extérieur des lois de chimie et physique."
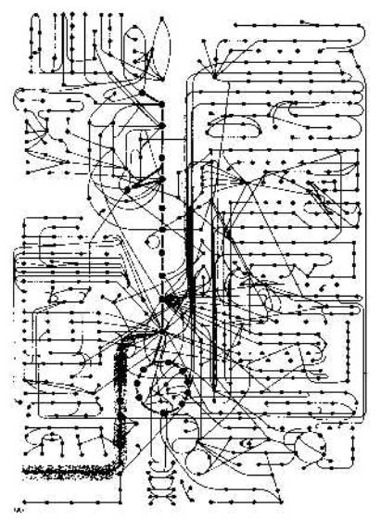
Mais professeur Bruce Alberts dit alors: "Quelque idée de comment
intriqué une cellule est, quand a envisagé comme une machine chimique, peut
être obtenu de Chiffre 2-35 (dans son livre). C'est un tableau qui en montre
seulement quelques-uns des chemins enzymatiques dans une cellule. Toutes ces
réactions se produisent dans une cellule qui est moins de 0.1 mm dans diamètre.
Et chacun exige une enzyme qui est elle-même le produit d'une série entière de
renseignements transfert et réactions de la protéine synthèse. Pour une petite
molécule typique - le serine de l'acide aminé, par exemple - il y a une
demi-douzaine ou plus d'enzymes qui peuvent le modifier dans chemin différant
chimiquement.
"Il peut être lié à AMP (adenylate), dans préparation pour synthèse de
la protéine, ou a dégradé à la glycine, ou a converti à pyruvate dans
préparation pour l'oxydation. Ce peut être des acetylate à acetyl CoA ou
transférer à un acide gras pour faire le serine du phosphatidyl. Tous ces
chemins différents rivalisent pour la même molécule du serine. Et les
compétitions semblables pour milliers d'autres petites molécules vont sur en
même temps. Un peut penser, que le système entier aurait besoin d'être
équilibré ainsi délicatement, que tout désordre mineur, tel qu'un changement
temporaire dans manger diététique, serait désastreux.
"En fait, la cellule est très écurie. Toutes les fois qu'il est
perturbé, la cellule réagit, donc comme restaurer son état initial. Il peut
adapter et continuer à fonctionner pendant privation de nourriture ou maladie.
Les mutations de beaucoup de genres peuvent éliminer des chemins de la réaction
particuliers. Et toujours - à condition, à ces certaines exigences minimums sont
satisfaites - la cellule survit. Il fait donc, parce qu'un réseau compliqué de
mécanismes du contrôle règle et coordonne le taux de ses réactions. ... Ici
nous nous inquiétons des mécanismes les plus simples qui règlent le courant de
petites molécules à travers les plusieurs chemins métaboliques seulement."
Alberts, B. et al. (1989:80).
Les chemins
métaboliques dans la cellule sont réglés. Comment?
Prof. Bruce Alberts: "Les concentrations des plusieurs petites molécules
dans une cellule sont amorties contre changements du majeur par un processus
connu comme règlement de la réaction. Il règle avec précision le flux de
métabolite à travers un chemin particulier, en augmenter temporairement ou
diminuant l'activité d'enzymes cruciales. Par exemple, la première enzyme d'une
série de réactions est inhibée par un effet de la réaction négatif du dernier
produit de ce chemin habituellement: Si les grandes quantités du dernier
produit accumulent, entrée supplémentaire de précurseurs dans le chemin de la
réaction est inhibée automatiquement. Où les chemins se branchent ou croisent,
comme ils font souvent, il y a des habituellement multiples points de contrôle
de produits définitifs différents. La complexité de tels processus du contrôle
de la réaction est illustrée dans Chiffre 2-37 (dans son livre). Il montre le
modèle de règlement de l'enzyme qui est observé dans un ensemble de chemins de
l'acide aminé apparentés.
"Le règlement de la réaction peut travailler presque instantanément et
être réversible; De plus, un produit de la fin donné peut activer des enzymes
qui mènent le long d'autres chemins aussi bien qu'inhibe des enzymes qui
causent sa propre synthèse." (1989:80).
La coordination du catabolisme (décomposition) et de la biosynthèse (rendant nouveau) dans la cellule. Après B. Alberts et autres (1989). Une personne intelligente a pensé dehors le schéma de circuit biochimique : le créateur du ciel et de la terre. Équiper l'a seulement trouvé et a compris. ¶
Comment fait le contrôle cellulaire bactérien ses alimentant réactions?
Pourquoi est-ce qu'il sait, lequel alimenter la réaction est juste et lequel
est faux?
Prof. F. C. Neidhardt et collègues: "Inhibition allosterique et de
l'activation jouer rôles importants dans régler le courant à travers alimenter
des chemins. Ici l'appareil simple de contrôle du produit de la fin du premier
ou un pas tôt dans sa formation ne peut pas appliquer. Le chemin de formation
des 12 métabilites du précurseur est mis en corrélation loin aussi
(quelques-uns sont cycliques) pour chacun, contrôler sa propre formation, en
inhibant un pas tôt unique. - Il n'y a pas de pas tôt uniques.
"Au lieu, les contrôles travaillent dans chacun des alimentant chemins
principaux intérieurement (Chiffre 4 dans son livre). Par exemple,
phosphoenolypyruvate, un intermédiaire de glycolyses, inhibe
phosphofructokinase, une enzyme qui catalyse une réaction trois pas dans le
même chemin en arrière. Et un ketoglutarate, un intermédiaire du tricarboxylique
cycle acide (TCA), inhibe synthéase du citrate qui catalyse la réaction deux pas
en arrière.
"De lui devrait être se souvenu, que pas tout le contrôle sur central
qui alimente des réactions peut être accompli par les interactions de l'allosterique.
Le renversement de courant dans l'Embden Meyerhof Parnas (EMP) chemin,
accomplir le gluconéogenèses pendant augmentation de cellules sur malate ou
pyruvate, par exemple, peuvent se produire, parce que tout, mais deux des
réactions entre pyruvate et phosphate de l'hexose, ayez des constantes de
l'équilibre loin de 1 pas. Et les deux, c'est pratiquement irrévocable, est mis
hors circuit par deux autres enzymes: phosphenolpyruvate synthetase qui met hors
circuit la kinase du pyruvate et une phosphatase spécifique qui hydrolzyes
fructose 1,6-diphosphate.
Equilibrer les taux de formation des 12 métabolites du précurseur à l'un
l'autre et aux demandes des chemins bio synthetiques, compliqué l'action de ces
composés comme effecteurs de l'allostérique comme illustré dans Chiffre 4
partialement. Mais le besoin pour coordination va au-delà le métabolite
carbone contenant. Alimenter des réactions doit fournir ATP et réduire un
nucléotide du pyridine (NADH2 et NADPH2). Ce n'est pas
surprenant, par conséquent, trouver, que ces composés sont des effecteurs de
l'allostérique à plusieurs points dans les alimentant chemins centraux."
(1990:310, 311).
Quand manger, quand le réparer, et quand le doubler, l'organisme
uniloculaire doit faire beaucoup de choses en même temps et dans le bon ordre.
Comment est-ce que cette créature minuscule fait cela?
Prof. F. C. Neidhardt et collègues affirment dans leur Physiology of the Bacterial Cell, sous le du titre "Règlement
de Gène Expression: Systèmes Multigène et Règlement Global":
"Beaucoup d'activités bactériennes impliquent la coordination du gène
à niveaux d'organisation qui est plus haute qu'unités du transcription al
individuelles. Le plus, si pas tout, les
opérons d'une cellule bactérienne appartiennent à plus haut niveau
organisations régulatrices que nous appellerons des RÉSEAUX RÉGULATEURS:
"L'opéron est un contrôle de la cellule du procaryotique. Cette mode
d'organisation - par lequel les gènes d'un chemin entier peuvent être unis
comme une unité du transcription al seule - accomplit une solution simple au
problème de gènes du co-régler de fonction apparentée. Pourquoi allez plus
loin? Est-ce qu'il y a un besoin pour la cellule bactérienne, déplacer au-delà
cette stratégie prospère évidente?
"Il y a au moins deux réponses à ces questions. Le premier est, que
quelques processus bactériens impliquent trop de gènes, être accommodé dans un
opéron réalisable seul. Cette situation est illustrée par la machinerie de la
traduction. Là un groupe d'au moins 150 produits du gène (ARN ribosomique,
protéines ribosomiques, initiation, allongement, et la terminaison compte,
synthéases de l'acyl-tARN aminé, et tARNs) est impliqué dans faire la protéine
d'acides aminés directement.
"Ils sont si nombreux, que coordonner leur synthèse en liant leurs
gènes dans un opéron seul, serait maladroit, si pas impossible. Toujours, le
règlement coordonné de ces gènes est important à l'efficacité totale
d'augmentation bactérienne. ... Cette coordination est accomplie, bien que ces
gènes soient organisés dans beaucoup de douzaines d'operons de l'non liant.
"La deuxième réponse est, que quelques processus bactériens impliquent
plusieurs gènes qui doivent être les deux réglés indépendamment et sujet à un
ravager, coordonner le contrôle. Cette situation est illustrée par l'ensemble
de gènes le plus clairement, en chiffrant des enzymes du catabolique qui sont
impliquées dans l'utilisation des sucres, les acides aminés, et les autres
composés pour carbone et énergie. Pendant augmentation dans un environnement
qui contient un mélange de tel compose, l'économie demanderait, que seulement
un substrat de la prime qui est capable le mieux satisfaire le carbone et
besoins de l'énergie de la cellule, devrait être métabolisé. (Pour les
bactéries entériques, le glucose est un tel substrat; Pour pseudo monads, le
succinate est.) Les opérons qui chiffrent des enzymes pour le métabolisme des
substrats secondaires, redondants devraient être réprimés. Cependant, chaque
opéron doivent être induits individuellement, quand son substrat analogue est
présent dans l'absence du substrat de la prime. Cette exigence double demande
un niveau d'organisation au-dessus de cela de l'opéron." (1990:351, 352).
Combien de systèmes du multi gène sont dans une cellule bactérienne là?
Comment est-ce qu'ils travaillent? Et comment complexe est-ce qu'ils sont?
Prof. F. C. Neidhardt et collègues: "Il est estimé, que la cellule
bactérienne a évolué plusieurs cent systèmes du multigène. Leur découverte et
l'analyse ont seulement seulement commencé. Quelques-uns des exemples bons
connus pour la bactérie entérique Escherichia
coli sont:
·
Réseaux qui sont impliqués en réponse à
limitation d'un ou un autre nutritif - carbone et sources de l'énergie, gaz
ammoniac, phosphate inorganique,
·
Réseaux qui sont impliqués dans réactions
de l'oxydation réduction et transport de l'électron
·
Réseaux qui s'inquiètent de réponse pour
endommager par oxydation, radiation, haute et basse température, et extrêmes de
pression osmotique
·
Réseaux divers, quelques-uns avec
sub-réseau qui provoque des changements du majeur dans la physiologie et
morphologie de la cellule.
La cellule bactérienne a évolué de divers chemins, tisser l'operons
individuel dans réseaux coordonnés; Les plusieurs mécanismes commencent à être
élucidé juste. Dans quelques cas, l'imaginez d'un régulateur de la protéine de
l'allosterique a été emprunté de règlement de l'opéron simplement: Un represseur
de la protéine ou l'activateur reconnaît une séquence particulière à qui est
commun le déterminant des régions de l'opérons du membre. Cela imagine est
utilisé dans le SOS, dégât de l'oxydation, et systèmes du transport de
l'électron anaérobies dans les bactéries entériques.
"Dans les autres systèmes le réseau est défini par un facteur du sigma
alternatif qui reprogramme le polymérise ARN pour reconnaître les organisateurs
des opérons du membre. Le choc de la chaleur et systèmes du sporulation de
plusieurs espèce bactérienne illustrent cette situation. Les autres réseaux
impliquent une combinaison de régulateurs de la protéine et sigma compte, comme
trouve dans le système de l'utilisation de l'azote de beaucoup de bactéries. Un
des réseaux les plus envahissant, les opérons stricts, est réglé par le
tetraphosphate du guanosine du nucléotide (ppGpp) dans une manière qu'encore
sera élucidée.
"Parce que beaucoup de réseaux régulateurs ont été reconnus comme
réponses cellulaires aux changements de l'environnement en premier, la
structure cérémonieuse d'un système de la stimulus-réponse peut fournir une
fonction de l'organisation utile pour la masse de renseignements vers chaque
réseau. Un système de la stimulus-réponse (Chiffre 1 dans son livre) contient
un chemin qui commence avec un stimulus de l'environnement qui affecte quelque
cible cellulaire ou sonde qui produit un signal. Ce système de la
stimulus-réponse directement - ou indirectement, quand le signal traverse un ou
plus de transducteurs - affecte l'activité ou synthèse d'un régulateur qui
contrôle la production. C'est quelque réponse adaptative au changement de
l'environnement habituellement. Ce système inclut souvent un mécanisme du
contrôle de la réaction qui autorise un retour à la condition du pré stimulus ou
à une nouvelle consonne de l'équilibre avec un environnement changé.
"Dans un réseau du gène bactérien, un stimulus de l'environnement (par
exemple, température changée, statut nutritif ou toxicité) serait représenté
comme affecter quelque sonde qui produirait un signal pour affecter l'activité
ou synthèse de (habituellement) un régulateur de la protéine des gènes du
membre. Les produits des gènes du membre exécuteraient quelque fonction(s)
cellulaire, encourager l'augmentation ou survie de la cellule. Les modulateurs
de quelque sorte permettraient un retour à l'état du pré stimulus d'expression
des opérons du membre ou établiraient un nouveau niveau de leur expression.
"C'est clair... que beaucoup de l'opérons d'E. coli est déjà assigné à un ou plus de réseaux. Avec l'aide de la structure, à condition dans Chiffre 1, nous en prouverons quelques-uns de ces réseaux en profondeur, illustrer des principes de physiologie du multigène et biologie moléculaire. Les premiers deux réseaux, être discuté, ont été choisis en partie, à cause de leur importance quantitative dans physiologie cellulaire. Ils contrôlent trois quarts de la capacité protéine synthétisant de la cellule bactérienne directement ou indirectement ensemble probablement." Neidhardt, F. C. (1990:352-357).
Répression Catabolique d'alimenter des gènes: Ce qui fait cette moyenne? Et
comment fait ce travail?
Prof. F. C. Neidhardt et collègues: "La plupart de l'espèce de
détachements du bactérien qu'un répertoire étendu de gènes qui chiffrent des
enzymes du catabolique a dirigé contre douzaines de composés différents de
diverse valeur alimentaire. Augmentation sur une source seule de carbone et
énergie - un substrat - exige un relativement haut niveau cellulaire des
enzymes qui métabolisent le substrat et nourrissent les produits du catabolique
dans les alimentant chemins centraux. Parce que chemins tout métaboliques dans
le courant cellulaire des métabolites qui ont été produits par ces enzymes du
catabolique.
"Les bactéries paraissent sentir l'à propos de chaque chemin du
catabolique dans une circonstance donnée et régler l'expression du courant du
gène en conséquence. Nous avons déjà noté, que le programme pour décider,
quelles enzymes faire et dans quelles quantités, doit être sophistiqué plus
qu'un, lequel simplement appels pour la cellule, induire des enzymes du
catabolique pour substrats qu'il a senti dans l'environnement. En effet, la
sophistication de ce programme cellulaire a dépassé la capacité de
physiologistes, comprendre ses complexités, en dépit d'un demi siècle d'effort
intensif.
"Les bactéries entériques emploient au moins quatre processus
différents, assurer le statut de la priorité de glucose comme un carbone et
source de l'énergie:
·
La
synthèse constitutive:
Les enzymes de catabolisme du glucose, est dans grande mesure une partie du
central EMP qui alimente le chemin. Ils sont produits à hauts niveaux, peu
importe ce que les substrats sont présents dans l'environnement.
·
L'exclusion
départ: Le Glucose
prévient l'entrée de certains autres induisant substrats par inactivation de
leur permeases.
·
La
répression transitoire:
Addition de produits alimentaires du glucose un voyageur, inhibition sévère de
synthèse d'enzymes du catabolique de l'départ pour vers demi d'une génération
en dépit de la présence de leur départ.
·
La
répression Catabolic:
Métabolisme de produits alimentaires du glucose une inhibition soutenue, mais
moins sévère de synthèse de l'enzyme provoquée, même dans la présence des
départs appropriés.
"Le non de ces processus est complètement compris. Et la répression du
catabolite est particulièrement complexe. Cependant, il inclut un élément vers
que beaucoup a été appris: l'a CAMP-CAP RÈSEAU. Dans E. coli l'opéron du lac
est un membre de ce réseau. Le nombre
total d'opérons du membre n'est pas su; mais ce peut être plusieurs douzaine et
inclure ceci, lesquels s'inquiètent de métabolisme de galactose, arabinose,
maltose, tryptophane, D-serine, et histidine. Les opérons du membre possèdent
une séquence du nucléotide caractéristique à qui la CAP de la protéine
régulatrice (protéine de l'activateur du gène du catabolique, produit du gène du crp) liens." Neidhardt, F. C. et al. (1990:357, 358).
Signalez la transduction dans systèmes du multi gène, ce qui fait cette
moyenne? Comment est-ce que cela travaille? Et comment complexe est-ce que
c'est?
Prof. F. C. Neidhardt et collègues: "Le réseau de l'arc avec les
autres trois réseaux, seulement a décrit, partagez un thème commun: Renseignements
qui traitent par interaction de la protéine-protéine. Laissez-nous considérer
ce sujet dans plus grande profondeur.
"Le traitement de renseignements physiologiques n'est pas un nouveau
sujet pour nous. Nous avons rencontré quelques aspects de renseignements
traiter, quand nous avons considéré les changements du conformational dans
certain (allostérique) protéines qui sont provoquées par complais réversible de
petites molécules spécifiques. Les petites molécules - lesquels sont produits
de la fin de chemins bio synthetiques, départs des chemins du catabolique, les
métabilite du précurseur ou le nucléotide - a transporté les renseignements
(vers leur concentration cellulaire) à la protéine de l'allosterique. Ce
processus a expliqué la plupart du contrôle sur activité de l'enzyme dans les
chemins bio synthétiques et dans alimenter des réactions. Et aussi pour
l'appareil stratégique central de physiologie de l'opéron - activation et
répression de transcription par protéines qui dans une conformation peuvent
lier ADN et dans un autre, ne pas." (1990:375-377).
Comment est-ce que la cellule bactérienne règle ses réseaux du multi gène?
Pourquoi est-ce que ces interactions sont importantes?
Prof. F. C. Neidhardt et collègues: Les interactions de la
protéine-protéine sont importantes dans régler les réseaux, décrit au-dessus.
Mais ils ne sont pas les moyens seuls de réseaux du multigène régulateurs. En
fait, les moyens moléculaires par que les groupes d'opérons différents sont
coordonnés dans leur expression, est assez divers.
Pour illustrer cette diversité, nous considérons trois réseaux qui
ressemblent à l'un l'autre dans un aspect utilitaire. Chacun est induit à
expression de haut niveau comme le résultat d'un changement du paramètre de
l'environnement seul, précis. Mais le mécanisme de l'énumération est très
différent dans chaque exemple.
"Un changement à haute température induit un ensemble d'ers 20 gènes
dans la plupart des cellules bactériennes. Cette énumération constitue la
réponse du chaleur-choc. C'est à une ampleur incroyable conservée
d'archaebactéries et eubactéries au-dessus à travers eucaryotes inférieur aux
plus hautes plantes et les animaux. Virtuellement chaque cellule microbienne et
chaque plante et la cellule animale répondent à un changement dans température.
À une gamme qui est juste précitée que qui est normal pour l'organisme donné en
induisant la synthèse de ses protéines du chaleur-choc.
"Un autre réseau - avec vers le même nombre de gènes - est le système
SOS. Ce groupe de gènes est induit à expression de haut niveau, quand le dégât
des expériences cellulaire à est ADN, comme par l'irradiation UV. Les produits
de la protéine de ces gènes sont impliqués dans plusieurs processus, ce dégât
de la réparation à ADN, et ce prévient les cellules de diviser, jusqu'à ce que
le tel dégât soit réparé. Ce réseau est induit par un décolleté du
proteolytique du represseur des opérons du membre.
"Le represseur est le produit du gène du lexA; Sur dégât à l'ADN d'une
cellule. La protéine RecA est activée, faciliter le décolleté du proteolytique
de LexA. Il induit les gènes LexA - Contrôlés qui constituent le système SOS de
cette façon.
"Encore une autre mode est illustrée par le réseau de l'oxyR. Ce
groupe d'une douzaine ou donc les gènes sont induits par oxidative endommagez
par qui est apporté sur par exemple, peroxyde de l'hydrogène. Les gènes du
membre chiffrent des enzymes protectrices, tel que catalase hydroperoxidase moi
et alkylhydroxyperoxidase. Ce réseau est induit par activation du régleur OxyR
positif, produit du gène de l'oxyR.
"Ces exemples servent, illustrer le fait que le règlement de réseaux du gène soit comme divers, comme est le règlement d'opérons individuel." Neidhardt, F. C. (1990:380, 381).
Pourquoi est-ce que la cellule vivante fait des protéines? Et pourquoi
est-ce qu'il sait, comment les faire? Et pourquoi est-ce qu'un certain ensemble
d'acides nucléiques traduits le texte dans protéines? En d'autres termes:
Pourquoi fait un certain signe (ou lettre) de l'ADN codez sachez, ce qu'il veut
dire? Et pourquoi est-ce qu'il veut dire ceci?
Bruce Alberts et collègues admettent dans leur manuel scolaire Molecluar Biology of the Cell
(1989:219), pourtant ils croient dans évolution: "Les processus
moléculaires sur que la synthèse de la protéine est basée, paraissez
inexplicablement complexe. Bien que nous puissions décrire beaucoup d'entre
eux, ils n'ont pas de sens conceptuel dans le chemin que la transcription ADN,
réparation ADN, et reproduction ADN fait. Comme nous avons vu, la synthèse de
la protéine dans actuels centres des organismes sur un très grand
ribonucleoprotein usine, le ribosome. Il consiste en protéines qui sont arrangées
autour d'un cœur de molécules du rARN.
"Pourquoi est-ce que les molécules du rARN devraient exister à tout?
Et comment est-ce qu'ils sont venus jouer une telle partie dominante dans la
structure et fonctionner du ribosome? La réponse nous aiderait, comprendre la
synthèse de la protéine, indubitablement. ... La synthèse de la protéine compte
aussi lourdement sur un grand nombre de protéines différentes qui sont liées au
rARNs dans un ribosome. La complexité d'un processus, avec tant de réagissant
réciproquement composants différents, a fait de grand nombre de biologistes
désespérer. Parce qu'ils ne croient pas, qu'ils seront jamais capables de
comprendre le chemin par que la synthèse de la protéine a évolué."
Nous pouvons nous souvenir encore, ce que professeur Bruce Alberts et ses
collègues ont réclamé dans leur manuel scolaire The Cell (1989:41): "Il n'y a aucune pièce dans la vue
contemporaine de vie... pour n'importe quoi autrement à l'extérieur des lois de
chimie et physique."
Est cette logique, est ce raisonnable, est-ce que ce scientifique est? Pas
à tout. La déclaration: "Il n'y a aucune pièce dans la vue contemporaine
de vie pour n'importe quoi à l'extérieur des lois de chimie et physique"
n'est pas logique, ce n'est pas raisonnable, ce n'est pas scientifique. La
raison: UNE loi vient toujours d'un législateur. Les lois de chimie et physique
contiennent des plus hauts renseignements scientifiques et des mathématiques.
Les scientifiques humains en bas ici ont compris jusqu'à maintenant seulement
quelques-uns d'eux sur terre. Quelques-unes de ces lois de chimie et physique
ils ont été capables de comprendre seulement un peu. Les renseignements et
mathématiques existent d'espèce humaine indépendamment. Le scientifique humain a
trouvé seulement et les comprises un petit. Les renseignements et mathématiques
sont quelque chose de non matériel, spirituel. Ils ont leur source dans un
monde spirituel, non matériel, dans le vrai Dieu de la Bible dont le nom est
Jéhovah.
Déjà sur le niveau du code nous pouvons savoir, qu'il a été étudié et été
fait par une personne intelligente. Un code (ou alphabet) est quelque chose de
spirituel, non matériel. Une personne intelligente doit l'étudier en premier.
Il doit sélectionner des certains signes en premier (lettres, etc.) et décide,
ce qu'ils signifieront. Avec ces signes, il peut écrire les mots, phrases,
chapitres, et le livre entier alors.
·
Les 20 acides aminés sont les 20 lettres
du code de l'acide aminé. Avec ces 20 lettres (et la grammaire a fourni avec
lui), la cellule vivante fait (écrit) ses protéines.
·
Les 4 acides nucléiques sont les 4 lettres
du code acide nucléique. Avec ces 4 lettres (et la grammaire a fourni avec
lui), la cellule vivante fait (écrit) son code génétique, sa chaîne ADN.
La première cellule de l'archal était sur terre de même que complexe
comme ceux, lesquels vivent maintenant. Il n'a pas "découvert" ou
"inventé" n'importe quoi. Même l'organisme uniloculaire le plus
"simple" est plus complexe, que n'importe quoi, quel homme a est
capable de faire labourer maintenant. L'ordinateur super le plus rapide
d'aujourd'hui est juste le jouet d'un enfant primitif, a comparé à la cellule
la plus simple qui vit aujourd'hui et lequel a vécu au commencement sur ce
monde, il y a quelque 3.8-4.0 milliards années.
L'hypothèse d'évolution ou néo-Darwinisme, comme maintenant a appris partout dans l'onde communément, n'a rien pour faire avec science sérieuse à tout. C'est une fausse religion. Il a ses racines dans les enseignements de Babylone ancien (Sumer Accad) et Egypte. Leurs prêtres païens ont déjà appris il y a quelques 3.500 à 4000 années, cette vie a évolué par elle-même de l'eau et boue de l'Euphrate et Tigre sur terre et du Nil. L'hypothèse moderne d'évolution ou néo-Darwinisme est une fraude énorme. Il cache sa vraie signification derrière la robe blanche de science moderne.