Perché sono cellule viventi? Perché sono capaci vivere? Perché la cellula è capace, fare i suoi propri fondamenti? Come complicato sono le sue reazioni chimiche? Se Lei, il caro lettore non capisce alcuni dei termini tecnici (o anche la maggior parte di loro), non si preoccupi di lui. In me voglio appena mostrarla così, quello che la piccola cellula che tutti conosce la biochimica, e quello che è capace fare. Poi chieda a Lei, per favore: perché questo piccolo organismo uni-cellula conosce tutti questi la biologia molecolare e la biochimica, quello che prima dove imparare un professore della biologia molecolare e la biochimica? Molti dei dettagli tecnici all'interno della cellula vivente sono complicato così, che gli esperti stessi sono stati capaci, ora capire solamente alcuni di loro. Alcuni loro sono stati capaci, capire solamente un poco, ed alcuni, non per niente. Almeno non fino a che ora.
Il metabolismo
Bruce Alberts è un Professore americano della Biologia. Lui è Presidente dell'Accademia Nazionale di Scienze (NAS) dell'America. Lui ed i suoi collaboratori credono che la vita sulla terra ha evoluto da sola. Loro uguagliano chieda nel loro manuale The Cell (La Cellula) (1989:41): "Non c'è stanza nella prospettiva contemporanea della vita... per qualsiasi cosa altro fuori delle leggi della chimica e la fisica."
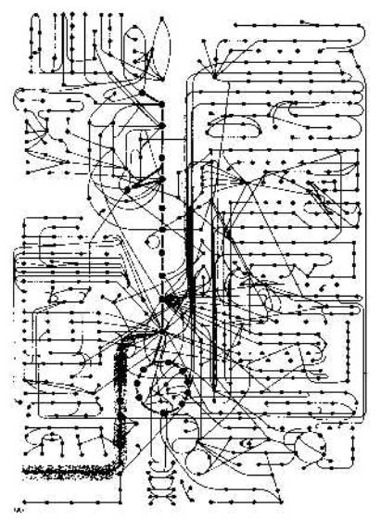
Ma dice poi Professor Bruce Alberts: "Dell'idea di come intricato è una cellula, quando vide come una macchina chimica, può essere ottenuto da Figura 2-35 (nel suo libro). È una tabella che mostra solamente alcuni dei sentieri enzimatici in una cellula. Tutte di queste reazioni accadono in una cellula che è meno che 0,1 mm in diametro. Ed ognuno richiede un enzima che si è il prodotto di una serie intera d'informazioni-trasferimento e reazioni della proteina-sintesi. Per una piccola molecola tipica - il serine d'acido d'amino, per esempio - c'è mezzo una dozzina o più enzimi che possono cambiarlo chimicamente in modo diverso.
"Può essere collegato ad Ampere (l'adenilato), in preparazione per sintesi della proteina, o degradò a glicine, o convertì a piruvato in preparazione per ossidazione. Può essere acetilate ad acetil CoA o può trasferire ad un acido grasso per fare fosfatidil serine. Tutti di questi sentieri diversi competono per la stessa molecola del serine. E le competizioni simili per migliaia d'altre piccole molecole seguono allo stesso tempo. Uno penserebbe, che il sistema intero avrebbe bisogno di essere bilanciato così eccellentemente, che qualsiasi sconvolgimento minore, come un cambio provvisorio in presa dietetica sarebbe disastroso.
"Infatti, la cellula è straordinariamente la stalla. Ogni qualvolta è perturbato, la cellula reagisce, così come ripristinare il suo stato iniziale. Può adattare e può continuare a funzionare durante fame o malattia. Mutazioni di qualche generi possono eliminare i particolari sentieri della reazione. Ed ancora - purché, quelli certi minimi requisiti sono soddisfatti - la cellula sopravvive. Fa così, perché una rete elaborata di meccanismi del controllo regola e coordina la percentuale delle sue reazioni. ... Qui noi concerniamo solamente coi più semplici meccanismi che regolano il flusso di piccole molecole attraverso i vari sentieri metabolici." Alberts, B. et al. (1989:80).
La coordinazione del catabolismo (analizzare) e della biosintesi (che rende nuovo) nella cellula. Dopo B. Alberts ed altri (1989). Lo schema circuitale biochimico si è pensato fuori da una persona intelligente: il creatore di cielo e di terra. Equipaggiare soltanto lo ha trovato e capito un piccolo.
Sentieri metabolici regolarono
I sentieri metabolici nella cellula sono regolati. Come?
Prof. Bruce Alberts: "Le concentrazioni delle varie piccole molecole in una cellula sono tamponate contro cambi del maggiore da un processo noto come regolamentazione della reazione. Fino accorda il flusso di metaboliti attraverso un particolare sentiero, aumentando temporaneamente o decrescendo l'attività d'enzimi cruciali. Per esempio, il primo enzima di una serie di reazioni di solito è interdetto da un effetto della reazione negativo del finale prodotto di quel sentiero: se le grandi quantità del finale prodotto accumulano, l'ulteriore entrata di precursori nel sentiero della reazione è interdetta automaticamente. Dove sentieri ramificano o s'intersecano, come spesso fanno loro, ci sono punti di solito multiplo di controllo di finale prodotto diverso. La complessità di tali processi di controllo di reazione è illustrata in Figura 2-37 (nel suo libro). Mostra il modello di regolamentazione dell'enzima che è osservato in un set di sentieri d'acido d'amino riferiti.
"Regolamentazione della reazione può funzionare quasi istantaneamente e può essere invertibile; in somma, un prodotto della fine data può attivare enzimi che conducono lungo gli altri sentieri così come interdica enzimi che provocano la sua sintesi." (1989:80).
Controlli d'Alimentare Reazioni
Come controlla le sue reazioni che alimentano la cellula batterica? Perché sa, qual alimentare reazione è giusto e qual è sbagliato?
Prof. F. C. Neidhardt e collaboratori: "L'inibizione d'allosterico e dramma d'attivazione gli importanti ruoli nel regolare il flusso attraverso alimentando sentieri. Qui la semplice apparecchiatura di controllo di prodotto di fine del primo o un primo passo nella sua formazione non può applicare. Il sentiero della formazione dei 12 metaboliti del precursore è collegato lontano anche (alcuni sono ciclici) per ognuno, controllare la sua formazione, interdicendo un primo passo unico. - Non ci sono primi passi unici.
"Invece, controlla lavori internamente in ognuno del principale alimentando sentieri (Figura 4 nel suo libro). Per esempio, fosfoenolipiruvate, un intermedio di glicolise, interdice fosfofructokinase, un enzima che catalizza indietro una reazione tre passi nello stesso sentiero. Ed un-ketoglutarate, un intermedio del ciclo d'acido tricarbossilico (TCA), interdice citrate sintase che catalizza indietro la reazione due passi.
"Dovrebbe essere ricordato, che non tutto il controllo su reazioni che alimentano centrali può essere completato da interazioni dell'allosterico. L'inversione di flusso nell'Embden-Meyerhof-Parnas (EMP) il sentiero, può accadere completare gluconeogenese durante la crescita di cellule su malate o piruvato, per esempio, perché tutti, ma due delle reazioni tra piruvato e fosfato dell'hexose, non abbiano lontano constante dell'equilibrio da 1. Ed i due, quello è praticamente irreversibile, sono aggirato da due altri enzimi: fosfenolpiruvatesintetase che aggira kinase del piruvato ed un fosfatase specifico che l'idrolizo fructose 1,6-difosfato.
Bilanciare le percentuali della formazione dei 12 metaboliti del precursore all'un altro ed alle richieste dei sentieri del biosintetico, coinvolto l'azione di questi combina come allosterico effectori, come illustrato parzialmente in Figura 4. Ma il bisogno per la coordinazione va oltre il metaboliti che carbone-contiene. Alimentando reazioni deve provvedere ATP e nucleotidi del piridine ridotto (NADH2 e NADPH2). Non è sorprendente, perciò, trovare, che questi combinano è allosterico effectori a molti punti nella centrale alimentando sentieri." (1990:310, 311).
Quando mangiando, quando riparandosi, e quando raddoppiandosi, l'organismo uni-cellula deve fare molte cose allo stesso tempo e nell'ordine destro. Come fa quella questa piccola creatura?
Prof. F. C. Neidhardt e collaboratori affermano nella loro manuale Fisiologia della Cellula Batterica sotto la Regolamentazione dell'intestazione "Dell'Expression di Gene: i Sistemi di Multigene e Regolamentazione Globale": "Molte attività batteriche comportano gene la coordinazione a livelli d'organizzazione che è più alta d'unità del trascrizione individuali. Di più, se non tutti, gli operoni di una cellula batterica appartengono ad organizzazioni di regulatoro di livello più alte di noi chiameremo le Reti di REGULATORIA:
"L'operon è un marchio di garanzia della cellula procariotica. Questa maniera d'organizzazione - da che cosa i geni di un sentiero intero possono essere uniti come una sola unità della trascrizione - realizza una semplice soluzione al problema di coregulato geni di funzione riferita. Perché vada ulteriore? C'è un bisogno per la cellula batterica, muoversi oltre questa strategia riuscito ed ovvia?
Ci sono almeno due risposte a queste domande. Il primo è, che alcuni processi batterici coinvolgono troppo molti geni, essere accomodato in un solo operon lavorabile. Questa situazione è illustrata dall'apparato della traduzione. Là un gruppo d'almeno 150 geni i prodotti (il ribosomale RNA, proteine del ribosomale iniziazione, l'allungamento, e fattori della terminazione amino-acil-tRNA il sintetasi, e tRNA) sono coinvolto direttamente nel fare proteina da acidi dell'amino.
"Loro sono così numerosi, che coordinare la loro sintesi collegando i loro geni in un solo operon, sarebbe goffo, se non impossibile. Ancora, regolamentazione coordinato di questi geni è importante all'efficienza complessiva della crescita batterica. ... Questa coordinazione è completata, anche se questi geni sono organizzati in molte dozzine di non-anello operoni.
"La seconda risposta è, che alcuni processi batterici comportano un numero di geni che devono essere ambo regolato indipendentemente e sottopone ad un avendo la priorità, mentre coordinando controllo. Questa situazione più chiaramente è illustrata dall'insieme di geni, mentre codificando enzimi cataboli che sono comportati nell'utilizzazione di zuccheri acidi dell'amino, ed altro combina per carbone ed energia. Durante la crescita in un ambiente della quale contiene una mistura tale combina, economia richiederebbe, che solamente un substrato del premio che è meglio capace per soddisfare il carbone ed energia ha bisogno della cellula, dovrebbe essere metabolizzato. (Per batteri enteri, glucosio è tale substrato; succinate è per pseudomonadi.) L'operoni che codifica enzimi per il metabolismo del secondario substrati ridondanti, dovrebbe essere represso. Ciascuni operon devono essere incitati individualmente in ogni modo, quando il suo substrato del cognato è presente nell'assenza del substrato del premio. Questo requisito duplice manda a chiamare un livello d'organizzazione sopra di quello dell'operon." (1990:351, 352).
Multigene regulatoro Reti
Quanti sistemi del multigene sono in una cellula batterica là? Come lavorano? E come complesso sono?
Prof. F. C. Neidhardt e collaboratori: "È valutato, che la cellula batterica ha evoluto molti cento sistemi del multigene. La loro scoperta e l'analisi ha cominciato solamente solo. Alcuni degli esempi buono-noti per il batterio enterio Escherichia coli è:
· Reti che sono comportate con risposta a limitazione di uno o un altro nutriente - carbone e fonti dell'energia, ammoniaca, fosfato inorganico
· Reti che sono comportate in reazioni dell'ossidazione-riduzione e trasporto dell'elettrone
· Reti che concernono con risposta per danneggiare da ossidazione, radiazione, temperatura alta e bassa, ed extremis di pressione dell'osmotico
· Reti miscellanee, alcuno con sub-reti che provocano cambi del maggiore nella fisiologia e la morfologia della cellula.
La cellula batterica ha evoluto modi diversi, tessere operoni individuale in reti coordinato; i vari meccanismi stanno cominciando solo ad essere delucidati. In dei casi, il concepisca di un moderatore di proteina d'allosterico è stato preso in prestito semplicemente da regolamentazione dell'operon: un repressor della proteina o attivatore riconosce una particolare sequenza alla qual è comune i controlli regioni dell'operoni del membro. Questo concepisce è usato nel SOS, danno d'ossidazione, e sistemi di trasporto d'elettrone anaerobi in batteri enteri.
"Negli altri sistemi la rete è definita da un fattore del sigma alternativo che il reprogrammi, il polimerase di RNA, riconoscere i promotori dell'operoni del membro. Il colpo di calore e sistemi della sporulazione di varia specie batterica illustrano questa situazione. Le altre reti comportano una combinazione di moderatori della proteina e fattori del sigma, come trovato nel sistema d'utilizzazione d'azoto di molti batteri. Una delle più reti del pervasive, l'operoni severo è regolato dal tetrafosfato di guanosine di nucleotide (il ppGpp) in una maniera che ancora sarà delucidata.
"Perché molte reti del regolatore furono riconosciute come risposte cellulari a cambi ambientali prima, la struttura formale di un sistema della stimolo-risposta può provvedere una funzione che organizza utile per la massa d'informazioni su ogni rete. Un sistema della stimolo-risposta (Figura 1 nel suo libro) contiene un sentiero che comincia con uno stimolo dall'ambiente che incide su dell'obiettivo cellulare o sensore che genera un segnale. Questo sistema della stimolo-risposta direttamente - o indirettamente, quando il segnale passa attraverso uno o più trasduceri - incide sull'attività o sintesi di un moderatore che controlla la produzione. Questa è della risposta adattabile al cambio ambientale di solito. Questo sistema include un meccanismo di controllo di reazione che permette un ritorno alla condizione del prestimulus spesso o ad un equilibrio nuovo consono con un ambiente cambiato.
"In un gene batterico la rete, uno stimolo dall'ambiente (per esempio, temperatura cambiata, status nutriente, o la tossicità) sarebbe dipinto come incidendo su del sensore che genererebbe un segnale, incidere sull'attività o sintesi di (solito) un moderatore della proteina del membro gene. I prodotti del membro geni compirebbero del funziona (cellulare), promuovere la crescita o sopravvivenza della cellula. Modulatori d'alcun genere abiliterebbero un ritorno allo stato del prestimulus d'espressione dell'operoni del membro o stabilirebbero un livello nuovo della loro espressione.
"È chiaro... che molto dell'operoni d'E. coli già sono assegnati ad uno o più le reti. Con l'aiuto della struttura, purché noi proveremo in Figura 1, alcuni di queste reti in profondità, illustrare principi della fisiologia del multigene e la biologia molecolare. Le prime due reti, essere discusso sono state scelte in parte, a causa della loro importanza quantitativa in fisiologia della cellula. Loro controllano direttamente o indirettamente insieme probabilmente tre-quarto della capacità che proteina-sintetizza della cellula batterica." Neidhardt, F. C. (1990:352-357).
Repressione di Catabolite di alimentare Geni
Repressione di Catabolite d'alimentare gene: quello che fa quel mezzo? E come fa quel lavoro?
Prof. F. C. Neidhardt e collaboratori: "Più specie di contee dei batteri che un repertorio esteso di geni che codificano enzimi cataboli diretto contro dozzine di diverso combina di valore nutrizionale e diverso. La crescita su una sola fonte di carbone ed energia - un substrato - richiede un relativamente livello cellulare ed alto degli enzimi che metabolizzano il substrato ed alimentano i prodotti cataboli nella centrale alimentando sentieri. Perché tutti i sentieri metabolici nella cellula fluiscono dal metaboliti che è stato prodotto da questi enzimi cataboli.
"Batteri sembrano sentire l'appropriatezza d'ogni sentiero catabolio in una circostanza data e regolare di conseguenza gene espressione del flusso. Noi già abbiamo notato, che il programma per decidere, che enzimi per fare ed in che quantità, deve essere sofisticato più che uno, quale manda a chiamare semplicemente la cellula, incitare enzimi cataboli per substrati che ha sentito nell'ambiente. Nonostante un mezzo-secolo di sforzo intensivo, la sofisticazione di questo programma cellulare ha ecceduto effettivamente, l'abilità di fisiologisti, capire le sue complessità.
"Batteri enteri assumono almeno quattro processi diversi, assicurare lo status prioritario di glucosio come un carbone e fonte dell'energia:
· Sintesi costitutiva: Gli enzimi di catabolismo di glucosio, è nella grande misura una parte dell'EMP sentiero che alimenta centrale. Loro sono prodotti a livelli alti, niente materia che substrati sono presenti nell'ambiente.
· Esclusione di Inducer: Glucosio previene l'entrata di certi altri substrati che incita da inactivazione del loro permeasi.
· Repressione transitoria: Somma di glucosio produce un'inibizione transitoria, severa di sintesi di inducibile enzimi cataboli per circa la metà di una generazione nonostante la presenza del loro inducer.
· Repressione catabolia: il metabolismo di glucosio produce un continuò, ma l'inibizione severa di sintesi dell'enzima indotta, anche nella presenza dell'induceri adatto.
"Non di questi processi è capito pienamente. E repressione del catabolite è particolarmente complessa. In ogni modo, include un elemento circa il qual è stato imparato molto: la Campo-berretto Rete. In E. coli il lac operon è un membro di questa rete. Il numero totale d'operoni del membro non è conosciuto; ma può essere molta dozzina e può includere quelli, quali concernono col metabolismo di galactose, arabinose, maltose, triptofane, D-serine, e histidine. Operoni del membro possiedono un nucleotide della caratteristica ordinano in sequenza a che il Berretto della proteina del regolatore (gene catabolismo proteina dell'activator, prodotto del crp gene) i legami." Neidhardt, T. F. C. et al. (1990:357, 358).
Segnali Trascondotta in Sistemi di Multigene
Segnali trascondotta in sistemi del multigene, quello che fa quel mezzo? Come funziona quello? E come complesso è?
Prof. F. C. Neidhardt e collaboratori: "La rete dell'arco con le altre tre reti, appena ha descritto, condivida un tema comune: informazioni che citano da interazione della proteina-proteina. Ci permetta di considerare questo tema nella più gran profondità.
"La lavorazione d'informazioni fisiologiche non è un tema nuovo per noi. Noi incontrammo degli aspetti d'informazioni citare, quando noi considerammo i cambi del conformazionale in certo (l'allosterico) proteine che sono provocate da complesso invertibile di piccole molecole specifiche. Le piccole molecole - quali sono prodotti della fine di sentieri del biosintetico, induceri di sentieri cataboli, metaboliti del precursore, o nucleotidi - portò informazioni (sulla loro concentrazione cellulare) alla proteina dell'allosterico. Questo processo incise per la maggior parte del controllo su attività dell'enzima in sentieri del biosintetico e nell'alimentare reazione. Ed anche per l'apparecchiatura strategica e centrale della fisiologia dell'operon - l'attivazione e repressione di trascrizione da proteine che in una conformazione possono legare DNA ed in un altro, non." (1990:375-377).
Reti di Multigene che regola
Come regola le sue reti del multigene la cellula batterica? Perché queste interazioni sono importanti?
Prof. F. C. Neidhardt e collaboratori: "Interazioni di Proteina-proteina sono importanti nel regolare le reti, descritto sopra di. Ma loro non sono il risuoli vuole dire di reti del multigene che regola. Infatti, il molecolare vuole dire da che gruppi d'operoni diverso sono coordinati nel loro expressio, è piuttosto diverso.
"Noi consideriamo che tre reti che assomigliano all'un altro in un aspetto funzionale illustrino questa diversità. Ognuno è incitato alto-livellare espressione come il risultato di un singola, cambio del parametro ambientale e bene-definito. Ma il meccanismo dell'insediamento è molto diverso in ogni esempio.
"Un turno a temperatura alta incita un set di circa 20 geni in più cellule batteriche. Questo insediamento costituisce la risposta della calore-colpo. È ad un'estensione incredibile conservata da archebatteri ed eubatteri su attraverso eucarioti più basso a piante più alte ed animali. Virtualmente ogni cellula microbica ed ogni pianta e cellula animale risponde un turno-su in temperatura. Ad un raggio d'azione che è solo su che è normale per l'organismo dato incitando la sintesi delle sue proteine del calore-colpo.
"Un'altra rete - con circa lo stesso numero di geni - è il sistema di SOS. Questo gruppo di geni è incitato per alto-livellare espressione, quando la cellula esperimenta danno ad è DNA, come da irradiazione d'UV. I prodotti della proteina di questi geni sono comportati nei vari processi che riparano danno a DNA e quello prevenga le cellule dal dividere, finché tale danno è riparato. Questa rete è incitata da una fenditura del proteolitico del repressore dell'operoni del membro.
"Il repressore è il prodotto del lexA gene; su danno al DNA di una cellula. La proteina di RecA è attivata, facilitare la fenditura del proteolitico di LexA. Incita con ciò i geni LexA-controllato che costituiscono il sistema di SOS.
"Ancora un'altra maniera è illustrata dalla rete dell'oxyR. Questo gruppo di una dozzina o così geni sono incitati da oxidative danneggi da che è portato su per esempio, perossido d'idrogeno. Il membro geni codificano enzimi protettivi, come catalase-hidroperoxidase io ed alkilidroxiperoxidase. Questa rete è incitata da attivazione del regolatore positivo OxyR, prodotto dell'oxyR gene.
"Questi esempi servono, illustrare che la regolamentazione di gene reti è come diverse, com'è la regolamentazione d'operoni individuale." Neidhardt, F. C. (1990:380, 381).
Sintesi della proteina, come sorto
Perché la cellula vivente fa proteine? E perché sa, come farli? E perché un certo set d'acidi nucleici traduce il testo in proteine? Nelle altre parole: Perché fa un certo segnale (o lettera) del DNA programmi sappia, quello che vuole dire? E perché vuole dire questo?
Bruce Alberts e collaboratori ammettono nel loro manuale la Molecular Biology of the Cell (Biologia Molecolare della Cellula) (1989:219), sebbene loro credono in evoluzione: "I processi molecolari sui quali è basata sintesi della proteina, sembri inesplicabilmente complesso. Anche se noi possiamo descrivere molti di loro, loro non hanno senso concettuale nel modo che la trascrizione di DNA, DNA ripara, ed il replicazione di DNA. Come abbiamo visto noi, sintesi della proteina in centri degli organismi attuali su una macchina del ribonucleoproteina molto grande, la ribosome. Consiste di proteine che sono sistemate circa un centro di molecole del rRNA.
"Perché molecole del rRNA dovrebbero esistere affatto? E come vennero a giocare tale parte dominante nella struttura e funzione della ribosome? La risposta ci aiuterebbe indubbiamente, capire sintesi della proteina. ... Sintesi della proteina conta anche pesantemente su un gran numero di proteine diverse che sono legate al rRNAi in un ribosome. La complessità di un processo, con così molti componenti che interagiscono diversi ha fatto molti biologi disperare. Perché loro non credono, che loro non saranno mai capaci di capire il sentiero dal quale evolve sintesi della proteina."
Noi ancora ricorderemmo, quello che Professor Bruce Alberts ed i suoi collaboratori chiesero nel loro manuale The Cell (La Cellula) (1989:41): non c'è stanza nella prospettiva contemporanea della vita... per qualsiasi cosa altro fuori delle leggi della chimica e la fisica."
È quello logico, è quello ragionevole, è quello scientifico? Non per niente. L'asserzione: "Non c'è stanza nella prospettiva contemporanea della vita per qualsiasi cosa fuori delle leggi della chimica e la fisica" non è logico, non è ragionevole, non è scientifico. La ragione: una legge viene da un legislatore sempre. Le leggi della chimica e la fisica contengono informazioni scientifiche e più alte e le matematiche. Scienziati umani hanno capito quaggiù sulla terra ora fino a che solamente alcuni di loro. Alcune di queste leggi della chimica e la fisica loro è stata capaci capire solamente un poco. Informazioni e le matematiche esistono indipendentemente d'umanità. Lo scienziato umano ha trovato solamente e li ha capiti un piccolo. Informazioni e le matematiche sono qualche cosa non-materiale, spirituale. Loro hanno la loro fonte in un mondo spirituale, non-materiale, nel vero Dio della Bibbia il cui nome è Geova.
Già sul livello del codice noi possiamo sapere, che si è pensato fuori ed è stato fatto da una persona intelligente. Un codice (o alfabeto) è qualche cosa spirituale, non-materiale. Una persona intelligente deve pensarlo fuori prima. Lui deve selezionare i certi segnali prima (le lettere, ecc.) e decide, quello che vorranno dire loro. Con questi segnali, lui può scrivere poi le parole, frasi, capitoli, ed il libro intero.
· I 20 acidi dell'amino sono le 20 lettere del codice d'acido d'amino. Con queste 20 lettere (e la grammatica approvvigionò con lui), la cellula vivente fa (scrive) le sue proteine.
· I 4 acidi nucleici sono le 4 lettere del codice d'acido nucleico. Con queste 4 lettere (e la grammatica approvvigionò con lui), la cellula vivente fa (scrive) il suo codice genetico, la sua catena di DNA.
La prima cellula dell'archea sulla terra era nel momento in cui complesso come quelli, quali ore stanno vivendo. Non ha "scoperto" o ha "inventato" qualsiasi cosa. Anche il più "semplice" organismo uni-cellula è più complessa, che qualsiasi cosa, quale uomo ha è capace fare ora coltivi. Il super-computer più veloce d'oggi è solo il giocattolo di un bambino primitivo, comparò alla più semplice cellula che sta vivendo oggi e quale ha vissuto all'inizio su questa terra, dei 3,8-4,0 miliardo anni fa.
L'ipotesi dell'evoluzione, o neo-Darwinismo, come ora insegnò comunemente in tutto il mondo, non ha niente per fare affatto con scienza seria. È una religione falsa. Ha le sue radici negli insegnamenti di Babilonia antica (Sumer Accad) e l'Egitto. I loro preti pagani già insegnarono dei 3.500 a 4.000 anni fa, quella vita sulla terra ha evoluto dall'acqua e fango dell'Eufrate e Tigri da sola e dal Nilo. L'ipotesi moderna dell'evoluzione o neo-Darwinismo è una frode enorme. Nasconde il suo vero significato dietro al vestito bianco di scienza moderna.