Batteri Gram-positivi e Gram-negativi. La loro membrana e muro. Come loro fanno i loro muri. Come loro differiscono. Cosa fu avuto bisogno, pensarli fuori e farli? Il loro muro della cellula prova, che loro tutti sono venuti da un antenato comune?
Prof. Frederic C. Neidhardt e collaboratori scrivono nella loro del manuale Physiology of the Bacterial Cell (Fisiologia della Cellula Batterica) (1990:25-27) sul muro dei batteri: "Batteri può essere diviso in due classi larghe - Gram-positivo e Gram-negativo. Batteri Gram-positivi e Gram-negativi differiscono in un numero d'importanti aspetti, oltre a struttura della busta. La distinzione Gram-positivo-Gram-negativa è un profondo. Non solo riflette una differenza fondamentale in struttura del muro, ma le altre importanti differenze biochimiche, fisiologiche, e genetiche come bene. La maggioranza, ma non tutti, batteri sono membri di questi due gruppi. Quelli quell'autunno fuori di questi gruppi è di due classi. Un gruppo è da vicino simile agli altri batteri, ma completamente muri della mancanza - il Mollicutes. L'altro gruppo, l'Archaebatteri produce muri che non contengono un murein tipico ed in molti altri rispetti differisca dalla cellula procariotica e tipica." (1990:25-27).
Muro della cellula
Scienziati ora hanno fondato fuori cosa sul muro della cellula? Come complesso è? Cosa deve uno tutti sappia di capire e farlo?
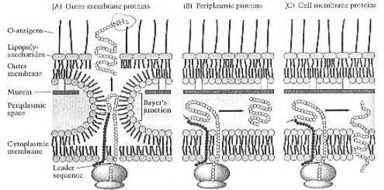
Prof. F. C. Neidhardt e collaboratori: "Come cellule vada, batteri sono particolarmente difficili; ovvero, loro sono duri rompere da meccanico vuole dire. E loro trattengono la loro forma le condizioni aspre addirittura sotto. Molta della durezza di batteri è riferita al loro muro della cellula che provvede appoggio meccanico e rigido e previene pressione dallo scoppiare le cellule (osmotico lisi). In somma, il muro della cellula rappresenta una difesa chimica e fisica contro chimici nocivo che può danneggiare la membrana della cellula. Batteri Gram-positivi e Gram-negativi differiscono considerevoli nella struttura dei loro muri della cellula, e nel ruolo di questa struttura, nell'affrontare con cambi ambientali. Alcuna porzione del muro d'occhiate delle cellule Gram-positive come una coperta spessa, quello di più cellule di Gram-negativo come un foglio fragile. In realtà, il muro Gram-negativo e sottile hanno anche forza del tensile considerevole." (1990:33).
Muro Gram-positivo
Prof. F. C. Neidhardt e collaboratori: "Il muro della cellula Gram-positivo consiste di un cappotto del multimolecular spesso di muerein (un tipo di peptidoglico) con minore ammontari d'altri polimeri, notevolmente acidi del teichoico; inframmezzò. La stoffa polimerica consiste di molti strati che sono avvolti circa la lunghezza ed ampiezza della cellula. Con ciò loro formano un sacco che determina la taglia e forma dell'organismo. I murein possono essere isolati intatti come una struttura, chiamò il murein sacculus. La forma del batterio dipende dalla forma del sacculus. La maggior parte di batteri sembra verghe (i bacilli), o eliche (lo spirocheti); alcun'occhiata come assi, stella di mare, o appiattì poliedra; ancora altri presumono forme più complesse, rese differente. ... Il murein rete rigido permette batteri, sopravvivere in media che sono comunemente ipotoni. Ovvero, loro hanno una pressione dell'osmotico più bassa, che quello del loro citoplasma. Nell'assenza di un rigido bustino-come struttura, la membrana scoppierebbe spingere contro, ed il lise delle cellule (= fluisca separatamente)." (1990:33-36).
Cellula Muro Gram-negativo e la Membrana esterna
Prof. F. C. Neidhardt e collaboratori: batteri "Gram-negativi hanno evoluto una soluzione radicalmente diversa al problema di proteggere la membrana citoplasmica. Il loro strato del murein molto è più sottile, che quello di batteri Gram-positivi, e loro fanno una struttura completamente diversa - una membrana esterna che è costruita su fuori di uno strato del murein sottile. La membrana esterna è chimicamente distinta dalle membrane biologiche e solite. È capace, resistere a danneggiare chimici. È una struttura due-strato, e la sua fogliolina interna assomiglia ad in composizione che della membrana citoplasma.
"La sua fogliolina esterna, d'altra parte ha un costituente unico nel luogo di fosfolipide. Questo componente è il lipopolisaccaride batterico, o LPS. Questa è una molecola complessa che non è trovata altrove in natura. Come un risultato, le foglioline di questa membrana sono molto asimmetriche. E le proprietà di quest'oppio pannicolo insolito differiscono notevolmente da quelli di una membrana biologica e regolare. L'abilità, escludere idrofobio combina, è insolito fra membrane biologiche.
"Esclusione d'idrofobio combina in batteri Gram-negativi, come in batteri Gram-positivi, è completato circondando le cellule con polisaccaride idrofobi. Ma questi differiscono in struttura ed organizzazione nei due gruppi. In ogni modo, i presenti membranosi ed esterni un vicolo cieco apparente: a causa della sua natura del lipide, può essere aspettaste escludere idrofilio combina come bene. In quel caso, nessuno combina, idrofobio o idrofilio, potrebbe attraversare la membrana esterna.
"Mentre risolvendo il problema di protezione della membrana citoplasma, batteri Gram-negativi sembrano avere creato un nuovo. Come trasportano i loro nutrienti questi organismi? Copiano nella membrana esterna le apparecchiature del trasporto attive della membrana citoplasma? Questa strategia non sola sarebbe un investimento rovinoso, ma anche potrebbe fare la membrana esterna nel momento in cui sensibile a sfide ambientali, com'è la membrana citoplasma. Ancora una volta, batteri hanno trovato una soluzione interessante: la membrana esterna ha canali speciali, quella licenza la diffusione passiva d'idrofilo combina come zuccheri, acidi dell'amino, ed i certi ioni. Questi canali consistono di molecole della proteina, porin adattamenti camati. Canali del porin sono stretti, solo la taglia destra, permettere l'entrata di combina più piccolo di 600-700 dalton (Figura 9 nel loro libro).
"I canali sono così piccoli, che idrofobo combinano entrerebbe in contatto col polare 'muro' del canale e con ciò sia escluso. A dell'estensione, E. coli possono selezionare porin con taglie del canale diverse, con risposta alle proprietà dell'osmotico del mezzo.
"Certo idrofilo combina, quale la cellula ha bisogno per sopravvivere, è più grande, che l'esclusione di porin. Queste grandi molecole includono vitamina B12, zuccheri che sono più grandi dei trisaccaride e celati di ferro. Tale combina croce la membrana esterna da meccanismi del permeazione separati, specifici. Loro utilizzano proteine che specialmente sono state disegnate a traslocate ognuno di questi combina. Cosi, la membrana esterna permette il passaggio di piccolo idrofilio combina, esclude idrofobo combina, grande o piccolo, e permette l'entrata di alcune più grandi molecole idrofile da meccanismi specialmente disegnati." (1990:37-40).
"La membrana esterna non è una struttura completamente separata; è connesso allo strato del murein e la membrana citoplasma. Il collegamento con lo strato del murein è interposto da due tipi d'interazioni, il più importante è un lipoproteina membranoso ed esterno. Questa proteina è presente in delle 700.000 copie per cellula che gli fabbrica la proteina più abbondante (numericamente, non da peso) in E coli.
Circa un terzo delle molecole del lipoproteina è covalente collegati insieme a murein e presa dell'aiuto le due strutture. L'altro tipo d'interazione è lo stretto (ma probabilmente non covalente) l'associazione di alcuni dei porin membranosi ed esterni con murein. Insieme, lipoproteini e porin provvedono in ogni cellula più di 400.000 contatti puntello-inoltri tra la membrana esterna ed il muro della cellula." (1990:41)
Presa elevare di Substrato specifici
Quale substrato specifico è il batterio E. coli capace prendere su? Come li prende su, in quali modi diversi?
Prof. F. C. Neidhardt e collaboratori: "Dei vari meccanismi del trasporto, pressoché tutti (inclusa diffusione facilitata, trasporto attivo e colpo-sensibile, e trasporto attivo e secondario) partecipi nella presa elevare di uno o un altro zucchero o alcol di zucchero da E. coli (Figura 4 nel loro libro). Interessatamente, il prendere di un particolare zucchera non segue un modello: per lactose dell'esempio è preso in E. coli da trasporto del protone, ma in Stafilococco aureus da un PTS.
"Acidi del amino sono trasportati da sistemi del trasporto attivi e colpo-sensibili o secondari. In E. coli, 14 sistemi del trasporto diversi che portano acidi dell'amino nella cellula. Molti modelli emergono da un esame degli amino acido trasporto sistemi di batteri. Alcuno del trasporto dei sistemi un gruppo d'acidi dell'amino con struttura simile. Spesso questi hanno sottosistemi per solamente uno degli acidi dell'amino. Questa ridondanza apparente serve uno scopo. Uno dei sistemi ha l'affinità alta (constante d'affinità 10 volte o più grande) e flusso alto. Ognuno ha vantaggi ovvi nei particolari ambienti. Oltre ad amino acido trasporto sistemi, E. coli hanno una varietà di sistemi del trasporto per cofactori, i vari ioni, ed intermediati metabolici." (1990:182, 183).
Come un Grammo negativo batterio fa le relative proprie proteine della busta. Dalla dei Neidhardt ed altri (1990:114) Fig. 8. La cellula molto piccola ha realizzato qui, che uomo non può ancora fare all'inizio di questo ventunesimo secolo: complessivo automatico, produzione automatica. Così quello, che ha pensato fuori ed ha fatto questo organismo di uni-cellula, deve conoscere molto più circa la scienza e la tecnologia, che tutto lo scienziato ora che vive su questa terra: il creatore, di cui il nome è Geova.
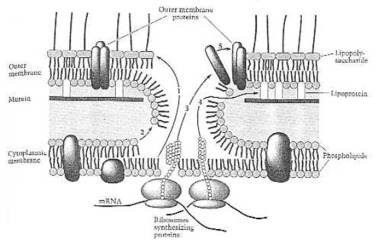
Reazione dell'Assemblea della membrana esterna. (1) Lipopolysaccharide; (2) fosfolipidi; (3) proteine esterne della membrana; (4) lipoproteina; (5) auto-complessivo della membrana esterna. Dalla fig. 14 dei Neidhardt ed altri (1990:121).
Regolamentazione d'Osmo e Porin Espressione
Come fa E. coli superati l'estremizzano di pressione dell'osmotico ambientale? E come fa i suoi porin, la legge attentamente nella sua membrana esterna?
Prof. F. C. Neidhardt e collaboratori: "Uno importante ambientale variabile con cui deve dare batteri, è l'osmolarità del loro ambiente. La cellula batterica utilizza molte strategie molecolari, contrattaccare gli effetti di extremis deleterio di pressione dell'osmotica ambientale.
"La membrana esterna è traversata da canali che sono fiancheggiati da proteine porin chiamati. In E. coli ci sono due porin del maggiore - OmpF ed OmpC. Loro tendono ad essere fatti come alternative all'un altro. La legge attentamente fatta da OmpC è lievemente più piccolo ed è fatto preferenzialmente a pressione dell'osmotica più alta e temperatura più alta. Questo modello vuole dire, quell'OmpC è la forma predominante durante la crescita di batteri enteri nel tratto intestinale. Ha il buon senso fisiologico per batteri enteri per avere più piccolo legge attentamente, quando risiedendo fuori nel tratto intestinale ed il più grande uni. Il più piccolo legge attentamente escluda molte delle molecole tossiche trovate nel tratto intestinale. Il più grande legge attentamente permetta substrati da diluisca soluzione, entrare la cellula legge attentamente prontamente, perché soluti devono diffondere attraverso la membrana esterna legge attentamente. E la percentuale di diffusione è proporzionale all'area croce-settoriale del legga attentamente, così come la differenza di concentrazione attraverso la membrana.
"I geni che codificano i porin notevoli così come certi altri geni che incidono su proprietà membranose ed esterne sono controllato da due proteine - prodotti dell'envZ collegato (per proteine della busta) ed ompR (per regolamentazione della proteina membranosa ed esterna) geni. La proteina d'EnvZ è una proteina membranosa e citoplasma che è pensata per funzionare come un osmo-sensore. Ha somiglianza strutturale e forte al certo kinases della proteina; sulla base di questa somiglianza e l'altra evidenza, ed investigatori credono dal fenotipo di sciocchezze e le altre mutazioni in envZ, quel EnvZ interagisce con la seconda proteina, OmpR. Incide sulla sua attività, da fosforilato esso.
"Proteina OmpR lega controcorrente specifica al DNA di promotori che controlla; il luogo vincolante è circa il -50 nel promotore dell'ompF, e -90 nel promotore dell'ompC. Un modello corrente, per com'espressione alternativa d'ompF è completato, è come segue (Figura 7 nel loro libro): pressione dell'osmotica alta conduce a fosforilazione d'EnvZ. E questo attivò proteina a turno fosforilate OmpR. Fosfo-OmpR (OmpR-P) reprime trascrizione d'ompF, legando ad un luogo tra il -40 ed il -60 nella regione di promotore d'ompF; attiva trascrizione dell'ompC, legando a luoghi di basso-affinità tra il -75 ed il -105 nella regione di promotore d'ompC. Quando la pressione dell'osmotica è bassa, il livello d'OmpR-P è basso. E perciò è incapace, attivare ompC; ma ancora è capace di attivare ompF, legando a luoghi d'alto-affinità nella -60 a -100 regione del promotore dell'ompF.
"Gli effetti d'OmpR-P, come uno stimolatore e come inibitore di trascrizione di questi due geni, è completato da un altro meccanismo. Il gene che micF è trascritto nella direzione opposta da ompC da un promotore, vicino quello d'ompC; micF codifica una 174-base RNA che è complementare ad una regione vicino l'inizio della comunicazione dell'ompF. La trascrizione del micF è stata mostrata, rendere impraticabile espressione d'ompC. Perché la regolamentazione d'espressione del micF è simile a quello d'ompC, le condizioni che incitano ompC inciteranno simultaneamente micF, e con ciò aiuta reprima ompF (Figura 7 nel loro libro).
C'è una rete di geni inoltre quelli per i due porin che sono regolati dal paio dell'envZ/OmpR di interagire proteine; alcuni di questi geni sono incitati, altri repressero. Il senso fisiologico e complessivo della scelta di geni che sono inclusi in questo sistema ancora è elusivo; l'importanza a noi della storia d'EnV/OmpR è, che ha provvisto un importante modello d'interazione della proteina-proteina in stimolo-risposta trasmette, incluso molti per che la funzione fisiologica è limpida." Neidhardt, R. C. et al. (1990:272, 373).
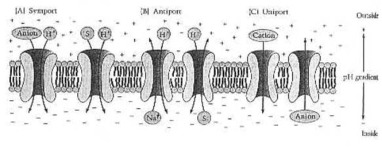
Sistemi secondari di trasporto attivo. (A) Reazioni di Symport. La pendenza di pH generata tramite gli azionamenti trasporto attivi primari (a sinistra) uno symport elettricamente neutro di un anione con un protone e (a destra) uno symport electrogenic di un soluto non caricare (s) insieme ad un protone.(B) Reazione di Antiport. La pendenza di pH guida (a sinistra) lo scambio elettricamente neutro di catione per un protone e (a destra) lo scambio elettronico di soluto non caricare (s) per un protone. (C) Reazioni di Uniport. La pendenza di pH guida un catione nella chiamata (di sinistra) di un anione fuori (di destra). Dalla dei Neidhardt ed altri (1990:178) Fig. 1
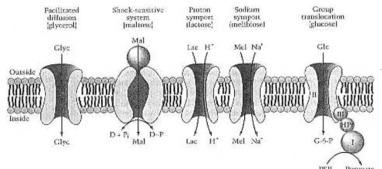
Meccanismi elemento mediati per l'assorbimento degli zuccheri e degli alcool di zucchero dal coly del E.. Il glicerolo (Glyc) entra tramite diffusione facilitata. Il maltosio (Mal) entra da un dipendente scuot sensibile del sistema su una proteina obbligatoria (sfera), su due proteine del trans-membrane (ovali) e su una proteina di energia transducing (freccia curva e doppio diretta) che idrolizza un donatore ad alta energia del fosfato (D~P). Il lattosio (Lc) entra dallo symport della proteina e dal melibiose (mel) da uno symport di Na+ di trasporto attivo secondario. Il glucosio (cromatografia gaseoliquido) entra da un I. T. con la mediazione degli enzimi in I (i), in HPr, nell'enzima II (ii) e nell'enzima III (iii); il prodotto intra cellulare del processo è glucosio 6-phosphate (G-6-p). Da Neidhardt ed altri (1990:182) Fig. 4. - Se, caro lettore, realmente non avete capito questo, non preoccuparsi. Ho desiderato appena mostrarlo, avete complicato questo siete e quanto quello, che ha pensato suo fuori, deve conoscere circa la scienza naturale, circa, la fisica e la biochimica.
Prima la Membrana, come sorto
Come è entrata la prima membrana in essendo? Potrebbe avere da solo airsen nella zuppa chimica e primordiale?
Prof. Siegried Scherer: "Fosfolipides è un principale-componente del biomembrani. Loro sono fatti da glicerin, acido del fosforico ed alifatico della lungo-catena combina. Loro sono congiunti insieme con glicerina come eteri o esters. Queste molecole, come tutte le altre sostanze che sono attive alla superficie (il tenside) abbia un idrofobo (= acqua-evitando), ed un idrofilo (= acqua-amoroso), componente. Molecole con queste struttura-caratteristiche sono capaci, venire insieme spontaneamente. Per esempio, come duplice strati, micelluli, e vescicole). Come tali sostanze sarebbero potute derivare le condizioni prebiotiche sotto, non è conosciuto.
"Membrane biologicamente attive, come noi li sappiamo da cellule viventi, sempre origini da membrane già esistenti. Non è sintetizzato loro de novo. I biomembrane non solo devono demarcare la cellula contro il suo dintorno. Ma deve anche, fra le altre cose, trasporti materiale attraverso la sua membrana. La costruzione complessa di specchi del biomembrani i compiti diversi delle membrane della cellula. Per esempio, il meccanismo di riconoscimento, cellula-cellula-comunicazione." (1998:146).
Hans Kleinig è un professore per la cellula-biologia all'Università di Freiburg. Uwe Maier è un professore per la cellula-biologia alla Philipps-Università Marburg, Germania. Loro scrivono nella loro del manuale Zellbiologie (Cellula biologia) (1999:73) sulla biosintesi di membrane: "Biomembrani non può derivare de novo nel Citoplasma di una cellula. I membrana-componenti, proteine, come lipidi sono sintetizzati nelle membrane loro di solito: in protociti: nella plasma-membrana..."
Risultato
Membrane biologicamente attive, come noi li sappiamo da cellule viventi, sempre origini da membrane già esistenti. Loro non sono sintetizzati di nuovo. Come tali sostanze sarebbero potute derivare le condizioni prebiotiche sotto, non è conosciuto
Noi ora abbiamo guardato brevemente a cellule Gram-positive e cellule Gram-negative. La cellula Gram-positiva ha un muro. La cellula Gram-negativa ha una membrana. Cellule Gram-positive, cellule Gram-negative, ed archaeabatteri differiscono molto dall'un l'altro. Loro non sono riferiti affatto all'un l'altro. Loro sono nel momento in cui diverso dall'un l'altro, come loro sono da piante ed animali. Ed uno non può tracciarli indietro ad un antenato comune e mitico. Già dei 3.5 a 3.8 miliardi anni fa, vicino l'inizio della storia di terra, c'erano cellule Gram-positive e Gram-negative, ed archebatteri, solo piaccia oggi.
Molte dei generi e specie, identificò a Warrawoona, NO Australia ancora sta vivendo oggi. Questi organismi uni-celluli erano nel momento in cui complesso, come quelli vivendo oggi. Loro non hanno evoluto a tutti nelle forme più alte della vita. Loro sono fabbriche chimiche e completamente automatiche, con un diametro di una piccola frazione di un millimetro. Come faccia fabbriche chimiche, case, coi loro muri e tetti sorgono oggi? Una casa (o fabbrica chimica) sorge nella mente di un essere umano ed intelligente per prima.
· Prima, lui ha un bisogno. È probabile che lui dica a lui: "Io voglio a lui abbia caldo ed asciuga, dove gli orsi e lupi non possono venire e possono uccidermi."
· Poi lui ha un augurio: "Io voglio avere una casa (o una tenda, o una caverna)."
· Poi lui fa un piano: "La mia casa sarà così e così grande. Avrà così e così molte stanze, porte, e finestre. Ed io lo farò con mattoni (o pietre, o tronchi)."
· Poi lui usa il suo piano della costruzione (la copia cianografica), energia, e costruendo materiali, e fabbrica la casa.
I primi tre passi che conducono alla casa sono puramente spirituali, non-materiali. Un essere umano ed intelligente, addestrato come un costruttore di casa li ha pensati fuori prima. Anche i mattoni nella casa (la loro forma e funzione), un essere umano ed intelligente ha pensato fuori prima. Gli atomi che costituiscono i mattoni (le pietre, e tronchi), non conoscano qualsiasi cosa una casa.
Materialisti hanno chiesto: tutto viene dalla materia. Ma una casa non scaturisce su dalle pietre o tronchi della taiga Siberiano, come un albero. Perché no? Perché il materiale dell'edificio della casa non conosce qualsiasi cosa la forma e funzione di una casa. Tutti che vengono da un essere umano ed intelligente che ha pensato a questo prima.
Gli atomi del muro (o membrana) del batterio sono al suo livello organizzativo e più basso. Loro sono solamente un mezzo ad una fine, come i mattoni nella casa. Un batterio molto è complicato più, che alcuna casa che mai ha costruito uomo. Uomo è capace di costruire una casa (ed una fabbrica chimica). Ma lui non è capace, fare cellula vivente, una piena fabbrica chimica automatizzato. Anche è complicato. È complicato più, che qualsiasi cosa, quale uomo mai ha fatto ora coltivi. Le prime cellule viventi su terra, dei 3,5-4 miliardi anni fa erano nel momento in cui complicato, come quelli vivendo oggi.
La cellula contiene sapere biochimico ed alto. Contiene informazioni e le matematiche. Ma la cellula stessa non ha scoperto o li ha inventati. Informazioni e le matematiche sono qualche cosa non-materiale, spirituale. Loro hanno la loro fonte in un mondo spirituale, non-materiale. Il batterio primo è dovuto essere pensato da adesso, anche fuori ed è dovuto essere fatto da qualcuno che conosce molto più scienza che alcun essere umano, ora vivendo su questa terra: il Creatore, il Dio della Bibbia il cui nome è Geova.