Capítulo 2: Biosíntese
Por
que é células vivas? Por que eles podem viver? Por que a célula é capaz, fazer
seus próprios blocos de edifício? Como complicado suas reacções químicas são?
Se você, querido leitor, não entende alguns dos termos técnicos (ou até mesmo a
maioria deles), não preocupe sobre isto. Em deste modo, eu há pouco quero
mostrar para você, isso que a célula minúscula que tudo sabem sobre bioquímica,
e o que pode fazer. Então o pergunte, por favor: Por que este organismo
unicelular pequeno sabe tudo isso sobre biologia molecular e bioquímica, o que
professor de biologia molecular e bioquímica teve que aprender primeiro? Muitos
dos detalhes técnicos dentro da célula viva são tão complicados, que os peritos
eles foram capazes, entender só alguns deles agora. Alguns eles foram capazes,
entender só um pouco, e alguns, não. Pelo menos não até agora.
Metabolismo
Bruce
Alberts é Professor americano de Biologia. Ele é o Presidente da Academia
Nacional de Ciências (NAS) de América. Ele e os colegas de trabalho dele
acreditam que vida em terra evoluiu por si só. Eles igualam reivindique no
livro de ensino deles The Cell (A Célula) (1989:41): "Não há nenhum
quarto na visão contemporânea de vida... para qualquer outra coisa fora das
leis de química e físicas."
Mas
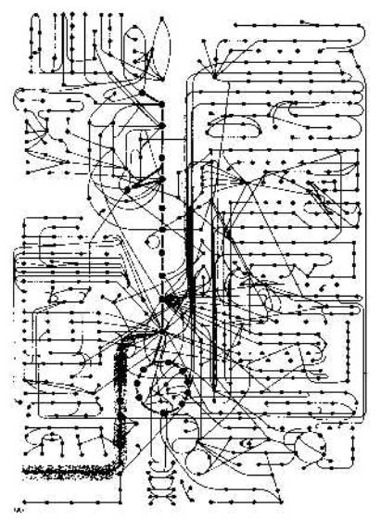
Professor Bruce Alberts diz então: "Algum ideia de como complicado uma
célula é, quando viu como uma máquina química, pode ser obtido de Figura 2-35
(no livro dele). É um quadro que mostra para só alguns dos atalhos enzimáticos
em uma célula. Todos estas reacções acontecem em uma célula que é menos de 0,1
mm em diâmetro. E cada requer uma enzima que se é o produto de uma série
inteira de informação transferência e reacções de proteína síntese. Para uma
molécula pequena típica - o serine de aminoácido, por exemplo - há meia dúzia
ou mais enzimas que podem modificar isto quimicamente de modo diferente.
"Pode
ser unido a AMP (adenilato, em preparação para síntese de proteína, ou degradou
a glicine, ou converteu a piruvate em preparação para oxidação. Pode ser
acetilate a acetil CoA ou pode transferir a um ácido gorduroso para fazer
serine de fosfatidil. Todos estes atalhos diferentes competem para a mesma
molécula de serine. E competições semelhantes para milhares de outras moléculas
pequenas vão ao mesmo tempo em. A pessoa poderia pensar, que o sistema inteiro
precisaria ser equilibrado assim finamente, que qualquer secundário transtorne,
como uma mudança temporária em entrada dietética, seria desastroso.
"Na
realidade, a célula é incrivelmente estável. Sempre que é perturbado, a célula
reage, para restabelecer seu estado inicial. Pode adaptar e pode continuar
funcionando durante fome ou doença. Mutações de muitos tipos podem eliminar
atalhos de reacção particulares. E ainda - contanto, para aquelas certas
exigências mínimas são satisfeitas - a célula sobrevive. Faz assim, porque uma
rede elaborada de mecanismos de controle regula e coordena a taxa de suas reacções.
... Aqui nós só nos preocupamos com os mecanismos mais simples que regulam o
fluxo de moléculas pequenas pelos vários atalhos metabólicos." Alberts, B. et al. (1989:80).
Atalhos metabólicos regularam
São
regulados os atalhos metabólicos na célula. Como?
Prof.
Bruce Alberts: "As concentrações das várias moléculas pequenas em uma
célula são buferas contra mudanças principais por um processo conhecido como
regulamento de avaliação. Bom afina o fluxo de metabolites por um atalho
particular, aumentando temporariamente ou diminuindo o actividade de enzimas
cruciais. Por exemplo, a primeira enzima de uma série de reacções normalmente é
inibida por um efeito de avaliação negativo do produto final daquele atalho: Se
quantidades grandes do produto final acumularem, entrada adicional de
precursores no atalho de reacção é inibida automaticamente. Onde atalhos se
ramificam ou cruzam, como fazem frequentemente eles, há pontos normalmente
múltiplos de controle de produtos finais diferentes. É ilustrada a complexidade
de tais processos de controle de avaliação em Figura 2-37 (no livro dele).
Mostra o padrão de regulamento de enzima que é observado em um jogo de atalhos
de aminoácido relacionados.
"Regulamento
de avaliação pode trabalhar quase instantaneamente e pode ser reversível; além,
um determinado produto de fim pode activar enzimas que conduzem ao longo de
outros atalhos como também iniba enzimas que causam sua própria síntese."
(1989:80).
A coordenação do catabolismo (que quebram para baixo) e do biosíntese (que faz novo) na pilha. Após B. Alberts et al. (1989). O diagrama de circuito bioquímico foi pensado para fora por uma pessoa inteligente: o criador do céu e da terra. Equipar somente encontrou-o e compreendeu um pouco. ¶
Controle de Abastecer Reacções
Como a
célula bactérial controla suas reacções abastecendo? Por que sabe, o qual
abastecendo reacção é certo e qual está errado?
Prof.
F. C. Neidhardt e colegas de trabalho: "Inibição de allosterico e jogo de
activação papéis importantes regulando o fluxo por abastecer atalhos. Aqui o
dispositivo simples de controle de produto de fim do primeiro ou um passo cedo
em sua formação não pode aplicar. O atalho de formação do 12 metabolites de
precursor é relacionado longe também (alguns são cíclicos) para cada, controlar
sua própria formação, inibindo um passo cedo sem igual. - Não há nenhum passo
cedo sem igual.
"Ao
invés, controla trabalhe interiormente em cada dos atalhos abastecendo
principais (Figura 4 no livro dele). Por exemplo, fosfoenolipiruvate, um
intermediário de glicolise, inibe fosfofructokinase, uma enzima que catalisa
uma reacção atrás três passos no mesmo atalho. E a-ketoglutarate, um
intermediário do tricarboxilico ciclo ácido (TCA), inibe sintase de citrate que
catalisa a reacção atrás dois passos.
"De
isto deveria se lembrar, que não todo o controle em reacções abastecendo
centrais pode ser realizado através de interacções de allostericos. A reversão
de fluxo no Embden-Meyerhof-Parnas (EMP) atalho, realizar gluconeogenesis
durante crescimento de células em malate ou piruvate, por exemplo, podem
acontecer, porque tudo, mas dois das reacções entre piruvate e fosfato de
hexose, não tenha constantes de equilíbrio longe de 1. E os dois, isso é
praticamente irreversível, é evitado através de duas outras enzimas: fosfenolpiruvatesintetase
que evita kinase de piruvate e um fosfatase específico que 1,6-difosfato de
frutose de hidrolse.
Equilibrar
as taxas de formação do 12 metabolites de precursor a um ao outro e para as
demandas dos atalhos biosintetios, envolvido a acção destas combinações como
efectuares de allostericos, como ilustrado parcialmente em Figura 4. Mas a
necessidade para coordenação vai além do metabolites carbono contendo.
Abastecendo reacções têm que prover ATP e nucleotides de piridine reduzido
(NADH2 e NADPH2). Então, não é surpreendente a achado,
que estas combinações são efectores de allostericos a vários pontos nos atalhos
abastecendo centrais." (1990:310, 311).
Ao
comer, ao se consertar, e ao se dobrar, o organismo unicelular tem que fazer
muitas coisas ao mesmo tempo e na ordem certa. Como esta criatura minúscula faz
isso?
Prof.
F. C. Neidhardt e colegas de trabalho declaram na Fisiologia de livro de ensino
deles da Physiology of the Bacterial Cell, do título, "Regulamento
de Expressão de Gene: Sistemas de Multigene e Regulamento Global":
"Muitos actividade bactérias envolvem coordenação de gene a níveis de
organização que é mais alta que unidades transcrever individuais. A
maioria, se não tudo, o operons de uma célula bacteriana pertencem a nível mais
alto organizações regulador que nós chamaremos REDES REGULADOR:
"O
operon é um carimbo oficial da célula procariotica. Este modo de organização -
por meio de que podem ser unidos os genes de um atalho inteiro como uma única
unidade transcrever - alcança uma solução simples ao problema de genes de
coregulato de função relacionada. Por que vá mais adiante? Há uma necessidade
pela célula bactérial, mover além desta estratégia próspera óbvia?
"Há
duas respostas pelo menos a estas perguntas. O primeiro é, que alguns processos
bactérial envolvem muitos genes, ser acomodado em um único operon executável.
Esta situação é ilustrada pela maquinaria de tradução. Lá um grupo de pelo
menos 150 produtos de gene (ribosomal RNA, proteínas de ribosomal, iniciação,
alongamento, e terminação fatora, sintetase de amino-acyl-tRNA, e tRNAs) é
directamente envolvido fazendo proteína de aminoácidos.
"Eles
são tão numerosos, que coordenar a síntese delas unindo os genes delas em um
único operon, seria desajeitado, se não impossível. Ainda, regulamento
coordenado destes genes é importante à eficiência global de crescimento
bactérial. ... Esta coordenação é realizada, embora estes genes sejam
organizados em muitas dúzias de operons não conectar.
"A
segunda resposta é, que alguns processos bactérias envolvem vários genes que
devem ser independentemente ambos regulados e sujeito a um anular, coordenando
controle. Esta situação é ilustrada claramente pelo conjunto de genes, enquanto
codificando enzimas catabolias que são envolvido na utilização de açúcares,
aminoácidos, e outras combinações para carbono e energia. Durante crescimento
em um ambiente que contém uma mistura de tal compõe, economia exigiria, que só
um substrato de prémio que é melhor capaz para satisfazer o carbono e energia
precisa da célula, deveria ser metabolizado. (Para bactérias enterias, glicose
é tal um substrato; para pseudomonades, é succinate.) Deveriam ser reprimidos o
operons que codificam enzimas para o metabolismo dos substratos secundários,
redundantes. Porém, cada operon devem ser induzidos individualmente, quando seu
substrato de cognato está presente na ausência do substrato de prémio. Estas
chamadas de exigência dobro para um nível de organização sobre o do
operon." (1990:351, 352).
Multigene Redes regulador
Quantos
sistemas de multigene estão lá em uma célula bactérial? Como eles trabalham? E
como complexo eles são?
Prof.
F. C. Neidhardt e colegas de trabalho: "É calculado, que a célula
bactérial evoluiu vários cem sistemas de multigene. A descoberta delas e
análise tem só começado. Alguns dos exemplos bom conhecidos para a bactéria
enteria Escherichia coli são:
·
Redes que
são envolvido com respeito a limitação de um ou outro nutriente - carbono e
fontes de energia, amónio, fosfato inorgânico,
·
Redes que
são envolvido em reacções de oxidacção reducção e transporte de eléctron
·
Redes que
estão preocupado com resposta danificar por oxidação, radiação, temperatura
alta e baixa, e extremes de pressão osmótica
·
Redes
misturadas, algum com subredes que provoca mudanças principais na fisiologia e
morfologia da célula.
A
célula bacterial evoluiu modos diversos, tecer operons individual em redes
coordenadas; os vários mecanismos há pouco estão começando a ser elucidados. Em
alguns casos, o legado de um regulador de proteína de allosterico foi pedido
emprestado simplesmente de regulamento de operon: um repressor de proteína ou
activador reconhece uma sucessão particular que está comum às regiões
controlando do operons de sócio. Este legado é usado no SOS, dano de oxidação,
e anaeróbio eléctron transporte sistemas em bactérias enterias.
"Em
outros sistemas a rede está definida por um factor de sigma alternativo que
reprograma polimerase de RNA reconhecer os promotores do operons de sócio. O
choque de calor e sistemas de sporulação de várias espécies bactérias
ilustram esta situação. Outras redes envolvem uma combinação de reguladores de
proteína e sigma factora, como acha no sistema de utilização de nitrogénio de
muitas bactérias. Um das redes mais penetrantes, o operons estrito, é regulado
pelo tetrafosfato de guanosine de nucleotide (ppGpp) até certo ponto que ainda
será elucidado.
"Porque
foram reconhecidas muitas redes regulador primeiro como respostas celulares para
mudanças ambientais, a estrutura formal de um sistema de estímulo resposta pode
prover uma função organizando útil para a massa de informação sobre cada rede.
Um sistema de estímulo resposta (Figura 1 no livro dele) contém um atalho que
começa com um estímulo do ambiente que afecta algum objectivo celular ou sensor
que gera um sinal. Este sistema de estímulo resposta directamente - ou
indirectamente, quando as passagens notáveis por um ou mais transducero -
afecta o actividade ou síntese de um regulador que controla a produção. Esta
normalmente é alguma resposta adaptável à mudança ambiental. Este sistema
inclui frequentemente um mecanismo de controle de avaliação que permite um
retorno à condição de prestimulus ou para uma consoante de equilíbrio nova com
um ambiente mudado.
"Em
uma rede de gene bactérial, um estímulo do ambiente (por exemplo, temperatura
mudada, estado nutriente, ou tóxico) seria descrito como afectando algum sensor
que geraria um sinal para afectar o actividade ou síntese de (normalmente) um
regulador de proteína dos genes de sócio. Os produtos dos genes de sócio
executariam algum funcção(s celular), promover o crescimento ou sobrevivência
da célula. Moduladores de algum tipo habilitariam um retorno ao estado de
prestimulus de expressão do operons de sócio ou estabeleceriam um nível novo da
expressão delas.
" Está claro... que muitos do operons de E. coli já são nomeados a um ou mais redes. Com ajuda do vigamento, contanto em Figura 1, nós provaremos alguns destas redes, ilustrar princípios de fisiologia de multigene e biologia molecular, a fundo. As primeiras duas redes, ser discutido, foram em parte escolhidas, por causa da importância quantitativa delas em fisiologia de célula. Junto eles controlam diretamente ou indirectamente provavelmente três quarto da capacidade proteína sintetizando da célula bactérial." Neidhardt, F. C. (1990:352-357).
Repressão de Catabolite de abastecer Genes
Repressão
de Catabolite de abastecer genes: o que faz aquele mau? E como faz aquele
trabalho?
Prof.
F. C. Neidhardt e colegas de trabalho: "A Maioria das espécies de
destacamentos de bactéria que um repertório extenso de genes que codificam
enzimas catabolias dirigiu contra dúzias de combinações diferentes de valor
nutricional diverso. Crescimento em uma única fonte de carbono e energia - um
substrato - requer um nível celular relativamente alto das enzimas que
metabolizam o substrato e alimentam os produtos catabolios nos atalhos
abastecendo centrais. Porque todos os atalhos metabólicos na célula fluem do
metabolites que foi produzido por estas enzimas catabolias.
"Bactérias
se aparecem sentir a conveniência de cada atalho catabolio em uma determinada
circunstância e regular expressão de fluxo de gene adequadamente. Nós já
notamos, que o programa por decidir, que enzimas para fazer e em que
quantidades, deve ser mais sofisticado que um, que pede a célula, induzir
enzimas catabolias para substratos que sentiu no ambiente, simplesmente.
Realmente, apesar de um meio século de esforço intensivo, a sofisticação deste
programa celular excedeu a habilidade de fisiólogos, entender suas
complexidades.
"Bactérias
enterias empregam quatro processos diferentes, assegurar o estado de prioridade
de glicose como um carbono e fonte de energia, pelo menos:
·
Síntese
constitutiva: As enzimas
de catabolismo de glicose, está em medida grande uma parte do EMP atalho
abastecendo central. Eles são produzidos a níveis altos, não importa que
substratos estão presentes no ambiente.
·
Exclusão
de Inducer: Glicose
previne a entrada de certos outros substratos induzindo por inactivação do
permeases delas.
·
Repressão
passageira: Adição de
glicose produz uma visitante, inibição severa de síntese de inducivel enzimas
catabolias para aproximadamente a metade de uma geração apesar da presença do
inducer deles.
·
Repressão
catabolia: Metabolismo de
glicose produz um continuou, mas inibição menos severa de síntese de enzima
induzida, até mesmo na presença do inducers apropriado.
"Não
destes processos são completamente compreendidos. E repressão de catabolite é
particularmente complexa. Porém, inclui um elemento sobre o qual muito foi
instruído: a REDE de CAMP-CAP. Em E. coli o operon de lac é
um sócio desta rede. O número total de operons de sócio não é conhecido;
mas pode ser vários dúzia e pode incluir esses, que estão preocupados com
metabolismo de galactose, arabinose, maltose, triptofan, D-serine, e histidine.
Operons de sócio possuem uma sucessão de nucleotide característica para qual o
CAP de proteína regulador (proteína de activador de gene catabolia, produto do
gene de crp) fitas." Neidhardt, F. C. et al. (1990:357,
358).
Tradução notável em Sistemas de Multigene
Tradução
notável em sistemas de multigene, o que faz aquele mau? Como isso trabalha? E
como complexo é?
Prof.
F. C. Neidhardt e colegas de trabalho: "A rede de arco com as outras três
redes, só descreveu, compartilhe um tema comum: processamento de informação
através de interação de proteína-proteína. Nos deixe considerar este tópico em
maior profundidade.
"O
processo de informação fisiológica não é um tópico novo para nós. Nós
encontramos alguns aspectos de processamento de informação, quando nós
consideramos as mudanças de conformacional dentro certo (allosterico) proteínas
que são provocadas por complexo reversível de moléculas pequenas específicas.
As moléculas pequenas - que são produtos de fim de atalhos biosintetios,
inducers de atalhos catabolios, metabolites de precursor, ou nucleotides -
carregou informação (sobre a concentração celular delas) para a proteína de
allosterica. Este processo respondeu pela maioria do controle em actividade de
enzima em atalhos biosintetios e abastecendo reacções. E também para o dispositivo
estratégico central de fisiologia de operon - activação e repressão de
transcrição por proteínas que em um conformação podem ligar o DNA e em outro,
não possa." (1990:375-377).
Redes de Multigene regulando
Como a
célula bacteriana regula suas redes de multigene? Por que estas interacções são
importantes?
Prof.
F. C. Neidhardt e colegas de trabalho: "Interacções de proteína-proteína
são importantes regulando as redes, descrito acima. Mas eles não são os meios
exclusivos de regular redes de multigene. Na realidade, os meios moleculares
pelos quais são coordenados grupos de operons diferente no expressio deles, é
bastante diverso.
"Nós
consideramos que três redes que se assemelham a um ao outro em um aspecto
funcional para ilustram esta diversidade. Cada é induzido para alto nivelar
expressão como o resultado de uma única, bem definido mudança de parâmetro
ambiental. Mas o mecanismo da indução é muito diferente em cada exemplo.
"Uma
troca para temperatura alta induz um jogo de cerca de 20 genes em a maioria das
células bactérias. Esta indução constitui a resposta de calor-choque.
É a uma extensão incrível conservada de arquebactérias e eubactéria para cima
por mais baixo eucariotes para plantas mais altas e animais. Virtualmente toda
célula microbiana e toda planta e célula animal responde um trocar para cima em
temperatura. Para uma gama que há pouco é anterior que é normal para o
determinado organismo induzindo a síntese de suas proteínas de calor choque.
"Outra
rede - com aproximadamente o mesmo número de genes - é o sistema de SOS. Este
grupo de genes é induzido para alto-nivelar expressão, quando a célula
experimenta dano para é o DNA, como através de irradiação de UV. Os produtos de
proteína destes genes são envolvidos em vários processos, aquele dano de
conserto para DNA, e aquele impeça para as células de dividir, até que tal dano
é consertado. Esta rede é induzida por uma divisão de proteolitico do repressor
do operons de sócio.
"O
repressor é o produto do gene de lexA; em dano para o DNA de uma célula.
A proteína de RecA é activada, facilitar a divisão de proteolitico de LexA.
Induz os genes LexA controlados que constituem o sistema de SOS assim.
"Ainda
outro modo é ilustrado pela rede de oxyR. Este grupo de uma dúzia de ou
assim genes são induzidos através de oxidative danifique por qual é trazido em
por exemplo, peroxide de hidrogénio. Os genes de sócio codificam enzimas
protetoras, como catalase-hidroperoxidase eu e alkilhidroxiperoxidase. Esta
rede é induzida por ativacção do regulador positivo OxyR, produto do gene de oxyR.
"Estes exemplos servem, ilustrar o facto que o regulamento de redes de gene é como diverso, como é o regulamento de operons individual." Neidhardt, F. C. (1990:380, 381).
Síntese de proteína, como surgido
Por
que a célula viva faz proteínas? E por que sabe, como os fazer? E por que um
certo jogo de ácidos nucleos traduz o texto em proteínas? Em outro palavra:
Por que faz um certo sinal (ou letra) do DNA codifique saiba, o que significa?
E por que significa isto?
Bruce
Alberts e colegas de trabalho admitem no livro de ensino deles Molecular
Biology of the Cell (Biologia Molecular da Célula) (1989:219), entretanto
eles acreditam em evolução: "Os processos moleculares nos quais síntese de
proteína é baseada, pareça complexo de inexplicável. Embora nós podemos
descrever muitos deles, eles não fazem sentido conceptual do modo que
transcrição de DNA, conserto de DNA, e replicação de DNA fazem. Como vimos nós,
síntese de proteína em centros de organismos atuais em uma máquina de
ribonucleoproteíne muito grande, o ribosomo. Consiste em proteínas que são
organizadas ao redor de um caroço de moléculas de rRNA.
"Por
que moléculas de rRNA deveriam existir nada? E como eles vieram fazer tal um
papel dominante na estrutura e função do ribosomo? A resposta nos, entender
síntese de proteína, ajudaria indubitavelmente. ... Síntese de proteína também
confia pesadamente em um número grande de proteínas diferentes que são ligadas
ao rRNAs em um ribosomo. A complexidade de um processo, com tantos componentes
interagindo diferentes, fez muitas biólogas desesperar. Porque eles não
acreditam, que eles sempre poderão entender o atalho pelo qual síntese de
proteína evoluiu ".
Nós
ainda poderíamos nos lembrar, o que Professor Bruce Alberts e os colegas de
trabalho dele reivindicaram no livro de ensino dele A Célula (1989:41):
não há nenhum quarto na visão contemporânea de vida... para qualquer outra
coisa fora das leis de química e físicas".
É
aquele lógico, é aquele razoável, é aquele científico? Não. A declaração: não
há nenhum quarto na visão contemporânea de vida por qualquer coisa fora das
leis de química e físicas " não é lógico, não é razoável, não é
científico. A razão: UMA lei sempre vem de um legislador. As leis de química e físicas
contêm informação científica mais alta e matemática. Cientistas humanos abaixo
aqui em terra entenderam até agora só alguns deles. Algumas destas leis de
química e físicas eles puderam entender só um pouco. Informação e matemáticas
existem independentemente de género humano. O cientista humano só achou e os
entendeu um pequeno. Informação e matemáticas são algo não material,
espiritual. Eles têm a fonte deles/delas em um espiritual, mundo não-materias,
no verdadeiro Deus da Bíblia cujo nome é Jeová.
Já no
nível do código nós podemos saber, que foi ideado e foi feito por uma pessoa
inteligente. Um código (ou alfabeto) é algo espiritual, não material. Uma
pessoa inteligente tem que idear isto primeiro. Ele tem que seleccionar certos
sinais primeiro (letras, etc.) e decide, o que eles quererão dizer. Com estes
sinais, ele pode escrever as palavras, orações, capítulos, e o livro inteiro
então.
·
Os 20
aminoácidos são as 20 letras do código de aminoácido. Com estas 20 letras (e a
gramática proveu com isto), a célula viva faz (escreve) suas proteínas.
·
Os 4 ácidos
nucleos são as 4 letras do código ácido nucleio. Com estas 4 letras (e a
gramática proveu com isto), a célula viva faz (escreve) seu código genético,
sua cadeia de DNA.
A
primeira célula de arqaea em terra era da mesma maneira que complexo como
esses, estão vivendo que agora. Não descobriu "ou "inventou"
qualquer coisa. Até mesmo o organismo unicelular mais simples " é mais
complexo, que qualquer coisa, qual homem tem pode fazer cultive agora. O super computador
mais rápido de hoje é há pouco o brinquedo de uma criança primitiva, comparou à
célula mais simples que está vivendo hoje e que se manteve nesta terra, uns
3,8-4,0 bilhões anos atrás, no princípio.
A hipótese de evolução, ou neo-Darwinismo, como agora comumente ensinou ao longo do mundo, não tem nada que ver com ciência séria nada. É uma falsa religião. Tem suas raízes nos ensinos de Babilónia antiga (Sumer-Accad) e Egito. Os padres pagãos deles já ensinaram uns 3.500 a 4.000 anos atrás, aquela vida em terra evoluiu por si só da água e lama do Eufrates e Tigris e do Nilo. A hipótese moderna de evolução ou neo-Darwinismo é um fraude enorme. Esconde seu verdadeiro significado atrás do vestido branco de ciência moderna.