Capítulo 6: Parede de Célula
Bactérias
Gram-positivas e Gram-negativas. A membrana delas e parede. Como eles fazem as
paredes delas. Como eles diferem. O que foi precisado, os idear e os fazer? A
parede de célula delas prova, que todos eles vieram de um antepassado comum?
Prof.
Frederic C. Neidhardt e colegas de trabalho escrevem na livro Physiology of
the Bacterial Cell (1990:25-27) sobre a parede das bactérias:
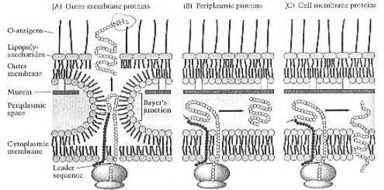
"Bactérias podem ser divididas em duas classes largas - Gram-positivo e
Gram-negativo. Bactérias Gram-positivas e Gram-negativas diferem em vários
aspectos importantes, além de estrutura de envelope. A distinção
Gram-positivo-Gram-negativa é uma profunda. Não só reflecte uma diferença
fundamental em estrutura de parede, mas outras diferenças bioquímicas,
fisiológicas, e genéticas importantes como bem. A maioria, mas não tudo,
bactérias, são os sócios destes dois grupos. Esse aquele outono fora destes
grupos é de duas classes. Um grupo é próximo semelhante a outras bactérias, mas
completamente paredes de falta - o Mollicutes. O outro grupo, o Arquebactérias,
produz paredes que não contêm um murein típico e em muitos outros cumprimentos
difira da célula procariotica típica." (1990:25-27).
Parede de célula
O que
descobriram os cientistas agora sobre a parede de célula? Como complexo é? O
que deve um tudo saiba em ordem entender e fazer isto?
Prof.
F. C. Neidhardt e colegas de trabalho: "Como células vá, bactérias são
particularmente duras; quer dizer, eles são difíceis de quebrar através de
meios mecânicos. E eles retêm a forma delas até mesmo debaixo de condições
severas. Muito da dureza de bactérias é relacionado à parede de célula delas
que provê apoio mecânico rígido e impede para pressão de estourar as células
(lise osmótico). Além, a parede de célula representa uma defesa química e
física contra substâncias químicas nocivas que podem prejudicar a membrana de
célula. Bactérias Gram-positivas e Gram-negativas diferem considerável na
estrutura das paredes de célula delas, e no papel desta estrutura, contendendo
com mudanças ambientais. Qualquer porção da parede de olhares de células
Gram-positivos como uma manta grossa, o da maioria das células Gram-negativas
como uma folha franzina. Até mesmo em realidade, a parede Gram-negativa magra
tem resistência à tração considerável." (1990:33).
Parede Gram-positiva
Prof.
F. C. Neidhardt e colegas de trabalho: "A parede de célula Gram-positiva
consiste em um casaco de multimolecular grosso de muerein (um tipo de peptidoglycan)
com menos quantias de outros polimer, notavelmente ácidos de teichoico;
entremeou. O tecido polímero consiste em muitas camadas que são embrulhadas ao
redor do comprimento e largura da célula. Assim eles formam um saco que
determina o tamanho e forma do organismo. O murein podem ser isolados intacto
como uma estrutura, chamou o sacculus de murein. A forma da bactéria
depende da forma do sacculus. A maioria do olhar de bactéria como varas (bacilos),
ou hélices (spirochetes); alguns olhar como fusos, estrela-do-mar, ou
aplainou polihedra; ainda outros assumem formas mais complexas, diferenciadas.
... A rede de murein rígida permite bactérias, sobreviver em mídia que são
comumente hipotonias. Quer dizer, eles têm uma mais baixa pressão osmótica, que
o do citoplasma delas. Na ausência de uma colete igual estrutura rígida, para
empurrar contra, estouraria a membrana e o lise de células (= flua separadamente)."
(1990:33-36).
Parede de Célula Gram-negativa e a Membrana
exterior
Prof.
F. C. Neidhardt e colegas de trabalho: bactérias "Gram-negativas evoluíram
uma solução radicalmente diferente ao problema de proteger a membrana
citoplasmia. A camada de murein delas está muito mais magra, que o de bactérias
Gram-positivas, e eles fazem uma estrutura completamente diferente - uma
membrana exterior que é construída fora de uma camada de murein magra. A
membrana exterior é quimicamente distinta das membranas biológicas habituais. É
capaz, resistir a substâncias químicas prejudiciais. É uma estrutura
dois estrata, e seu folheto interno está em composição que da membrana
citoplasmia.
"Por
outro lado, seu folheto exterior tem um componente sem igual no lugar de fosfolipido.
Este componente é o lipopolisaccaride bacteriano, ou LPS. Esta é uma molécula
complexa que não é achada em outro lugar em natureza. Como resultado, os
folhetos desta membrana são extremamente assimétricos. E as propriedades deste
bi-camada incomum diferem consideravelmente desses de uma membrana biológica
regular. A habilidade, excluir combinações hidrofóbicas, é incomum entre
membranas biológicas.
"Exclusão
de combinações hidrofóbicas em bactérias Gram-negativas, como em bactérias
Gram-positivas, é realizado cercando as células com polisaccarides hidrofóbico.
Mas estes diferem em estrutura e organização nos dois grupos. Porém, a membrana
exterior apresenta um impasse aparente: pode ser esperado que exclua
combinações hidrófilas como bem por causa de sua natureza de lipídio. Naquele
caso, nenhuma combinação, hidrofóbico ou hidrófilo, poderia cruzar a membrana
exterior.
"Enquanto
resolvendo o problema de protecção da membrana citoplasma, bactérias
Gram-negativas parecem ter criado um novo. Como estes organismos transportam os
nutritivas delas? Eles copiam na membrana exterior os dispositivos de
transporte activos da membrana citoplasma? Esta estratégia não só seria um
investimento esbanjador, mas também poderia fazer a membrana exterior da mesma
maneira que sensível a desafios ambientais, como é a membrana citoplasma. Uma
vez mais, bactérias acharam uma solução interessante: A membrana exterior tem
canais especiais, aquela licença a difusão passiva de combinações hidrófilas
como açúcares, aminoácidos, e certos íones. Estes canais consistem em moléculas
de proteína, porin habilmente chamados. Canais de porin são estreitos, só o
tamanho certo, permitir a entrada de combinações menor que 600-700 dalton
(Figura 9 no livro deles).
"Os
canais são tão pequenos, que combinações hidrofóbicas entrariam em contacto com
o polar 'parede' do canal e assim seja excluído. Até certo ponto, E. coli
podem seleccionar porin com tamanhos de canal diferentes, com respeito às
propriedades osmóticas do médio.
"Certas
combinações hidrófilas das que a célula precisa para sobreviver, é maior, que a
exclusão de porin. Estas moléculas grandes incluem vitamina B12, açúcares que
são maiores que trisacarido e chelates férreo. Tal compõe cruz a membrana
exterior através de mecanismos de permeação separados, específicos. Eles
utilizam proteínas que especialmente foram projetadas a translocate cada destas
combinações. Assim, a membrana exterior permite a passagem de combinações
hidrófilas pequenas, exclui combinações hidrofóbicas, grande ou pequeno, e
permite a entrada de algumas moléculas hidrófilas maiores através de mecanismos
especialmente projetados." (1990:37-40).
"A
membrana exterior não é uma estrutura completamente separada; é conectado à
camada de murein e a membrana citoplasma. A conexão com a camada de murein é
mediada por dois tipos de interacções, o mais importante é um lipoprotéine de
membrana exterior. Esta proteína está presente em uns 700.000 cópias por célula
que faz isto a proteína mais abundante (numérico, não através de peso) em E
coli.
"Sobre
um terço das moléculas de lipoprotéine são covalento unidos junto a murein e
cabo de ajuda as duas estruturas. O outro tipo de interacção é o apertado (mas
provavelmente não covalento) associação de alguns dos porin de membrana
exteriores com murein. Junto, lipoprotéine e porin provêem em cada célula mais
de 400.000 contactos escora semelhantes entre a membrana exterior e a parede de
célula." (1990:41).
Acolhimento de Substratos específicos
Quais
substratos específicos são a bactéria E. coli capaz levar para cima?
Como os leva, de quais modos diferentes?
Prof.
F. C. Neidhardt e colegas de trabalho: "Dos vários mecanismos de
transporte, quase tudo (incluindo difusão facilitada, transporte activo
choque sensível, e transporte activo secundário) participe no acolhimento de um
ou outro açúcar ou álcool de açúcar por E. coli (Figura 4 no livro
deles). De forma interessante, o acolhimento de um açúcar particular não segue
um padrão: por exemplo é levada lactose em E. coli através de transporte
de próton, mas em Staphylococcus aureus por um PTS.
Ácidos
de "Amino são transportados através de sistemas de transporte activos
choque sensíveis ou secundários. Em E. coli, 14 sistemas de transporte
diferentes que trazem aminoácidos na célula. De um exame dos sistemas de
transporte de aminoácido de bactérias, emergem vários padrões. Algum do
transporte de sistemas um grupo de aminoácidos com estrutura semelhante.
Frequentemente estes têm subsistema para único dos aminoácidos. Esta redundância
aparente serve um propósito. Um dos sistemas tem afinidade alta (afinidade 10
vezes constantes ou maior) e fluxo alto. Cada tem vantagens óbvias em
particular ambientes. Além de sistemas de transporte de aminoácido, E. - coli
tem uma variedade de sistemas de transporte por cofactors, vários íones, e
intermediários metabólicos." (1990:182, 183).
Como uma Grama-negativo bactéria faz suas próprias proteínas do envelope. De Neidhardt et de Fig. 8 do al. (1990:114). A pilha minúscula conseguiu aqui, que homem não pode ainda fazer no começo deste século XXI: conjunto automático, produção automática. Assim esse, que pensou para fora e fêz este organismo do unicelular, deve saber muito mais sobre a ciência e a tecnologia, do que todo o cientista que vive agora nesta terra: o criador, cujo o nome é Jeová.
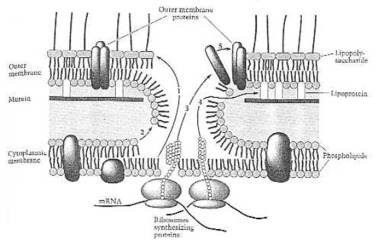
Reação do conjunto da membrana exterior. (1) Lipopolysaccharide; (2) phospholipids; (3) proteínas exteriores da membrana; (4) lipoprotein; (5) self-conjunto da membrana exterior. De Neidhardt et al.de Fig. 14 do (1990:121).
Regulamento de Osmo e Expressão de Porin
Como
faz E. coli superado o extremes de pressão osmótica ambiental? E como
faz seus porin, os poros em sua membrana exterior?
Prof.
F. C. Neidhardt e colegas de trabalho: "Uma variável ambiental importante,
com que bactérias têm que negociar, é o osmolaridade do ambiente delas. A
célula bacterista utiliza várias estratégias moleculares, contrariar os efeitos
de extremes danoso de pressão osmótica ambiental.
"A
membrana exterior é atravessada por canais que estão forrado através de
proteínas porin chamados. Em E. coli há dois porin principais -
OmpF e OmpC. Eles tendem ser feitos como alternativas a um ao outro. Os
poros feitos por OmpC são ligeiramente menores e são feitos preferencialmente a
pressão osmótica mais alta e temperatura mais alta. Este padrão significa,
aquele OmpC é a forma predominante durante crescimento de bactérias enterias na
área intestinal. Faz sentido fisiológico bom por bactérias enterias terem poros
menores, ao residir na área intestinal e maior fora. Os poros menores excluem
muitas das moléculas tóxicas achados na área intestinal. Os poros maiores
permitem substratos de dilui solução, entrar no poro de célula prontamente,
porque solutas têm que difundir pelos poros de membrana exteriores. E a taxa de
difusão é proporcional à área cruz secional do poro, como também a diferença de
concentração pela membrana.
"Os
genes que codificam os porin principais como também certos outros genes que
afectam propriedades de membrana exteriores são controlados através de duas
proteínas - produtos do envZ unido (para proteínas de envelope) e
ompR (para regulamento de proteína de membrana exterior) genes. A
proteína de EnvZ é uma proteína de membrana citoplasma que é pensada para
funcionar como um osmo sensor. Tem semelhança estrutural forte a certo kinases
de proteína; em base desta semelhança e outra evidência, e do fenótipo de
tolice e outras mutações em envZ, acreditam os investigadores, aquele EnvZ
interage com a segunda proteína, OmpR. Afecta seu actividade, fosforilate isto.
"Proteína
OmpR liga especifico rio acima ao DNA de promotores que controla; o local que
liga é ao redor -50 no promotor de ompF, e -90 no promotor de ompC.
Um modelo actual, para como expressão alternativa de ompF é realizada, é
como segue (Figura 7 no livro deles/delas): pressão osmótica alta conduz a
fosforilação de EnvZ. E isto ativou em troca proteína fosforilate OmpR.
Fosfo-OmpR (OmpR-P) reprime transcrição de ompF, ligando para um
local entre -40 e -60 na região de promotor de ompF; activa transcrição de ompC,
ligando para locais de baixo afinidade entre -75 e -105 na região de promotor
de ompC. Quando a pressão osmótica for baixa, o nível de OmpR-P é baixo.
E então é incapaz, activar ompC; mas ainda pode activar ompF,
ligando para locais de alto afinidade na -60 a -100 região do promotor de ompF.
"Os
efeitos de OmpR-P, como um stimulator e como inhibitor de transcrição destes
dois genes, é completado por outro mecanismo. O micF de gene é
transcrito na direcção oposta de ompC de um promotor, perto do de ompC;
micF codifica um 174-base RNA que é complementar a uma região perto do
começo da mensagem de ompF. A cópia de micF foi mostrada,
bloquear expressão de ompC. Porque o regulamento de expressão de micF
é semelhante a isso de ompC, as condições que induzem ompC
induzirão micF simultaneamente, e assim ajuda reprime ompF
(Figura 7 no livro deles).
Há uma
rede de genes além desses para os dois porin que são regulados pelo par de
envZ/OmpR de proteínas interagindo; alguns destes genes são induzidos, outros
reprimiram. O senso fisiológico global da escolha de genes que são incluídos
neste sistema ainda é enganoso; a importância para nós da história de EnV/OmpR
é, que proveu um padrão importante de interacção de proteína-proteína em redes
de estímulo resposta, enquanto incluindo muitos para os quais a função
fisiológica é cristalina claro.". Neidhardt, R. C. et al. (1990:272,
373).
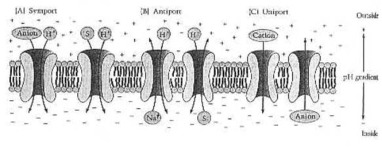
Sistemas secundários do transporte ativo. (A) Reações de Symport. O gradient do pH criado por movimentações do transporte ativo preliminares (na esquerda) um symport electroneutral de um anion com um proton, e (na direita) um symport electrogenic de um solute não carregar (S) junto com um proton. (B) Reação de Antiport. O gradient do pH dirige (na esquerda) a troca electroneutral de um cation para um proton e (na direita) a troca eletrônica de um solute não carregar (S) para um proton. (C) Reações de Uniport. O gradient do pH dirige um cation na chamada (esquerda) de um anion para fora (direito). De Neidhardt et al. de Fig.1 do (1990:178).
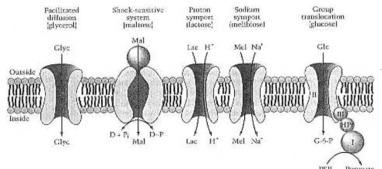
Mecanismos ortador mediados para o recepção dos açúcares e dos álcoois de açúcar pelodo E. coli . O glycerol (Glyc) entra pela difusão facilitada. O maltose (Mal) entra por um dependente choc-sensível do sistema em uma proteína obrigatória (esfera), em duas proteínas do transmembrane (ovals), e em uma proteína da energia-transducing (seta curvada, dobro-dirigida) que hydrolyzes um doador high-energy do phosphate (D~P). O lactose (Lc) entra pelo symport da proteína e pelo melibiose (mel) por um symport de Na+ do transporte ativo secundário.O glucose (glc) entra por um Técnico Especialista de Filial com o mediation dos enzymes em I (i), em HPr, em enzyme II (ii), e em enzyme III (iii); o produto intracelular do processo é o glucose 6-phosphate (G-6-p). De Neidhardt et al. de (1990:182) Fig.4. - se você, caro leitor, não compreender realmente este, não se preocupar. Eu quis apenas mostrá-lo, você complicou este é, e quanto esse, que pensou de para fora, tem que saber sobre a ciência natural, aproximadamente, a física e o bioquímico. ¶
Primeiro Membrana, como surgido
Como a
primeira membrana entrou em ser? Poderia ter airsen por si só na sopa química
primordial?
Prof.
Siegried Scherer: "Fosfolipides são um principal-componente dos
biomembrana. Eles são feitos de glycerin, ácido de fosforico e combinações de
alifaticos de longo cadeia. Eles são unidos junto com glicerina como éteres ou
esters. Estas moléculas, como todas as outras substâncias que são activo à
superfície (tenside) tenha um hidrofóbico (= água evitando), e um hidrófilo (=
água amoroso), componente. Moléculas com estes estrutura característica são capazes,
vir junto espontaneamente. Por exemplo, como dobrar camadas, micelules, e
vesículas). Como tais substâncias pudessem ter surgido debaixo de condições
prebiotias, não é conhecido.
"Membranas
biologicamente activas, como nós os conhecemos de células vivas, sempre
origine de já membranas existentes. Eles não são sintetizados nenhum novo de de
(em cima de novamente, novamente). O biomembrana não só tem que demarcar a
célula contra seus ambientes. Mas também deve, entre outras coisas, material de
transporte por sua membrana. A construção complexa de espelhos de biomembrana
as tarefas diversas das membranas de célula. Por exemplo, mecanismo de
reconhecimento, célula-célula comunicação." (1998:146).
Hans
Kleinig é um professor para célula biologia ao Universidade de Freiburg. Uwe
Maier é um professor para célula biologia ao Philipps-Universidade Marburg,
Alemanha. Eles escrevem na Zellbiologie (Célula-biologia) de livro de
ensino deles (1999:73) sobre o biosíntese de membranas: "Biomembrana não
podem surgir nenhum novo de no Citoplasma de uma célula. Os
membrana componentes, normalmente são sintetizadas proteínas, como lipídios,
nas membranas eles: em protocites: na protoplasma membrana..."
Resultado
Membranas
biológicas activas, como nós os conhecemos de células vivas, sempre origine
de já membranas existentes. Eles não são sintetizados novamente. Como tais
substâncias pudessem ter surgido debaixo de condições prebiotias, não é
conhecido
Nós
olhamos agora brevemente para células Gram-positivas e células Gram-negativas.
A célula Gram-positiva tem uma parede. A célula Gram-negativa tem uma membrana.
Células Gram-positivas, células Gram-negativas, e arquebactéria diferem muito
de um ao outro. Eles não são relacionados a um ao outro nada. Eles são da mesma
maneira que diferente de um ao outro, como eles são de plantas e animais. E a
pessoa não os pode localizar atrás a um antepassado comum mítico. Já uns 3,5 a
3,8 bilhões anos atrás, se aproxime o começo da história de terra, havia
células Gram-positivas e Gram-negativas, e arquebactérias, há pouco igual
hoje.
Muitas
do genera e espécies, identificados a Warrawoona, NO a Austrália, ainda está
vivendo hoje. Estes organismos unicelulares eram da mesma maneira que complexo,
como esse vivo hoje. Eles não evoluíram nada em formas mais altas de vida. Elas
são fábricas químicas completamente automáticas, com um diâmetro de uma fracção
minúscula de um milímetro. Como faça fábricas químicas, casas, com as paredes
dela e telhados, surgem hoje? Uma casa (ou fábrica química) surge no princípio
na mente de um ser humano inteligente.
·
Primeiro,
ele tem uma necessidade. Ele poderia dizer a ele: "Eu quero a isto
tenha morno e seca, onde os ursos e lobos não podem vir e podem me matar."
·
Então ele
tem um desejo: "Eu quero ter uma casa (ou uma barraca, ou uma
caverna)."
·
Então ele
faz um plano: "Minha casa será fulano de tal grande. Terá fulano de tal
muitos quartos, portas, e janelas. E eu farei isto com tijolos (ou pedras, ou
troncos)."
·
Então ele
usa o plano de construção dele (fotocópia azul), energia, e construindo
materiais, e faz a casa.
Os
primeiros três passos que conduzem para a casa são puramente espirituais,
não matérias. Um ser humano inteligente, treinado como um construtor de casa,
os ideou primeiro. Até mesmo os tijolos na casa (a forma delas e função), um
ser humano inteligente ideou primeiro. Os átomos que compõem os tijolos
(pedras, e troncos), não saiba nada sobre uma casa.
Materialistas
reivindicaram: Tudo vem de assunto. Mas uma casa não cresce para cima fora das
pedras ou tronco do taiga sibíria, como uma árvore faz. Por que não? Porque o
material de edifício da casa não sabe nada sobre a forma e função de uma casa.
Tudo aquilo vem primeiro de um ser humano inteligente que pensou nisto.
Os
átomos da parede (ou membrana) da bactéria está a seu mais baixo nível
organizacional. Eles são só uns meios a um fim, como os tijolos na casa. Uma
bactéria é complicada muito mais, que qualquer casa que o homem alguma vez
construiu. Homem pode construir uma casa (e uma fábrica química). Mas ele não é
capaz, fazer célula viva, uma fábrica de substância química automatizada cheia.
É muito complicado. É mais complicado, que qualquer coisa, qual homem sempre
fez agora gaveta. As primeiras células vivas em terra, uns 3,5-4 bilhões anos
atrás, eram da mesma maneira que complicado, como esse vivo hoje.
A célula contém experiência bioquímica alta. Contém informação e matemática. Mas a própria célula não descobriu ou os inventou. Informação e matemáticas são algo não matérial, espiritual. Eles têm a fonte delas em um espiritual, mundo não matérial. Consequentemente, também a bactéria deve ter sido ideada primeiro e deve ter sido feita por alguém, que sabe muito mais sobre ciência, que qualquer ser humano, se mantendo nesta terra agora: o Criador, o Deus da Bíblia cujo nome é Jeová.