Capítulo 2: La Biosíntesis
¿Por qué es células vivientes? ¿Por qué ellos pueden vivir? ¿Por qué la célula es capaz, hacer sus propios ladrillos? ¿Cómo complicadas sus reacciones químicas son? Si usted, estimado lector, no entiende algunas de las condiciones técnicas (o incluso la mayoría de ellos), no se preocupe por él. De esta manera, yo quiero apenas mostrarlo, eso que la célula diminuta que todos saben sobre la bioquímica, y lo que puede hacer. ¿Entonces pregúntelo, por favor: Por qué este organismo del una-célula pequeño sabe todos esto sobre la biología molecular y bioquímica, lo que profesor de biología molecular y bioquímica tenía que aprender primero? Muchos de los detalles técnicos dentro de la célula viviente son tan complicados, que los expertos ellos han sido capaz, para entender ahora sólo unos de ellos. Algunos ellos han sido capaces, para sólo entender un poco el pedazo, y algunos, nada. Por lo menos no hasta ahora.
Bruce Alberts es Profesor americano de Biología. Él es Presidente de la Academia Nacional de Ciencias (NAS) de América. Él y sus co-obreros creen que la vida en la tierra ha evolucionado solo. Ellos igualan exige en su libro de texto The Cell (La Célula) (1989:41): "No hay ningún cuarto en la vista contemporánea de vida... para nada más fuera de las leyes de química y físicas."
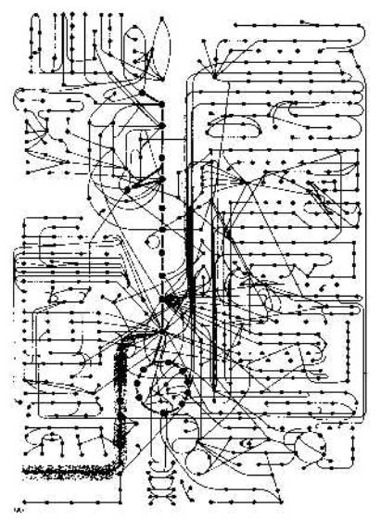
Pero Profesor Bruce Alberts dice entonces: "Alguna idea de cómo intrincado una célula es, cuando vio como una máquina química, puede obtenerse de Figura 2-35 (en su libro). Es un mapa que muestra sólo algunas de los sendas enzimáticos en una célula. Todos que estas reacciones ocurren en una célula que está menos de 0.1 mm en el diámetro. Y cada uno requiere una enzima que se es el producto de una serie entera de información-traslado y reacciones de la proteína-síntesis. Para una molécula pequeña típica - el serine del aminoácido, por ejemplo - hay mitad una docena o más enzimas que pueden modificarlo químicamente de la manera diferente.
"Puede unirse al AMP (adenylate), en la preparación para la síntesis de la proteína, o degradó a la glicina, o convirtió al piruvate en la preparación para la oxidación. Puede ser los acetylate al acetilo CoA o puede transferir a un ácido graso para hacer los serine del fosfatidil. Todos que estas sendas diferentes compiten para la misma molécula del serine. Y las competiciones similares para los miles de otras moléculas pequeñas siguen al mismo tiempo. Uno podría pensar, que el sistema entero necesitaría ser equilibrado así finamente, que cualquier menor disgustado, como un cambio temporal en la succión dietética, sería desastroso.
"De hecho, la célula es increíblemente estable. Siempre que sea perturbador, la célula reacciona, para restaurar su estado inicial. Puede adaptar y puede continuar funcionando durante inanición o enfermedad. Las mutaciones de muchos tipos pueden eliminar las sendas de la reacción particular. Y todavía - con tal de que, ese ciertos requisitos mínimos se reúnen - la célula sobrevive. Hace para que, porque una red detallada de mecanismos del mando regula y coordina la proporción de sus reacciones. ... Aquí nosotros sólo nos preocupamos por los mecanismos más sencillos que regulan el flujo de moléculas pequeñas a través de las varias sendas metabólicas." Alberts, B. et al. (1989:80).
La coordinación del catabolismo (subdivisión) y de la biosíntesis (que hace nuevo) en la célula. Después de B. Alberts et al. (1989). El esquema circular bioquímico ha sido pensado hacia fuera por una persona inteligente: el creador del cielo y de la tierra. Servir lo ha encontrado y ha entendido solamente un poco.
Se regulan las sendas metabólicas en la célula. ¿Cómo?
Prof. Bruce Alberts: "Las concentraciones de las varias moléculas pequeñas en una célula son los buffer contra los cambios del comandante por un proceso conocido como la regulación de la regeneración. Multa-pone a punto el flujo de metabolitos a través de una senda particular, aumentando temporalmente o disminuyendo la actividad de enzimas cruciales. Por ejemplo, la primera enzima de una serie de reacciones normalmente se inhibe por un efecto de la regeneración negativo del último producto de esa senda: Si las cantidades grandes del último producto aumentan, la entrada extensa de precursores en la senda de la reacción se inhibe automáticamente. Donde las sendas echan ramas o cortan, cuando ellos hacen a menudo, hay puntos normalmente múltiples de mando de productos finales diferentes. Se ilustra la complejidad de tales procesos de mando de regeneración en Figura 2-37 (en su libro). Muestra el modelo de regulación de la enzima que se observa en un juego de sendas del aminoácido relacionadas.
"La regulación de la regeneración puede trabajar casi instantáneamente y puede ser reversible; además, un producto del extremo dado puede activar enzimas que llevan a lo largo de otras sendas así como inhiba enzimas que causan su propia síntesis." (1989:80).
¿Cómo hace el mando celular bacteriano sus reacciones alimentando? ¿Por qué sabe, qué alimentando la reacción es correcto y cuál está equivocado?
Prof. F. C. Neidhardt y co-obreros: "La inhibición Allosterica y obra de activación los papeles importantes regulando el flujo a través de alimentar las sendas. Aquí el dispositivo sencillo de mando de producto de extremo del primero o un paso temprano en su formación no puede aplicar. La senda de formación de los 12 metabolitos del precursor sé interrelaciona lejos también (algunos son cíclicos) para cada uno, controlar su propia formación, inhibiendo un único paso temprano. - No hay ningún único paso temprano.
"En cambio, los mandos trabajan internamente en cada uno de las sendas alimentando principales (Figura 4 en su libro). Por ejemplo, el fosfoenolipiruvate, un intermedio de glicólisis, inhibe fosfofructocinase, una enzima que catalizar una reacción atrás tres pasos en la misma senda. Y un-ketoglutarate, un intermedio del tricarboxilo el ciclo ácido (TCA), inhibe sintase del citrate que catalizar la reacción atrás dos pasos.
"Debe recordarse, que no todo el mando en las reacciones alimentando centrales puede lograrse por las interacciones allostericas. La inversión de flujo en el Embden-Meyerhof-Parnas (EMP) la senda, lograr el gluconeogenesis durante el crecimiento de células en malate o piruvate, por ejemplo, pueden ocurrir, porque todos, pero dos de las reacciones entre el piruvate y fosfato del hexose, no tenga las constantes de equilibrio lejos de 1. Y los dos, eso es prácticamente irreversible, se desvía por dos otras enzimas: fosfenolpiruvatesintetase que desvía el kinase del piruvate y un fosfatase específico que el hidrolzies fructosa 1,6-diphosfato.
"Para equilibrar las proporciones de formación de los 12 metabolites del precursor entre si a y a las demandas de las sendas del biosintetico, envuelto la acción de estos compuestos como el effectores allostericos, como ilustrado parcialmente en Figura 4. Pero la necesidad para la coordinación va más allá del metabolitos carbono-conteniendo. Alimentando las reacciones deben proporcionar ATP y el nucleótidos del piridina reducido (NADH2 y NADPH2). Por consiguiente, no es sorprendente al hallazgo, que estos compuestos están los efectores allostericos en varios puntos en las sendas alimentando centrales." (1990:310, 311).
Al comer, al repararse, y cuando se dobla, el organismo del una-célula debe hacer muchas cosas al mismo tiempo y en el orden correcto. ¿Cómo esta criatura diminuta hace eso?
Prof. F. C. Neidhardt y co-obreros declaran en su libro de texto de la Physiology of the Bacterial Cell (Fisiología de la Célula Bacteriana) bajo el título "La Regulación de Expresión del Gen: Los Sistemas de Multigene y la Regulación Global":
"Muchas actividades bacterianas involucran la coordinación del gen a los niveles de organización que es más alta que las unidades de la trascripción individuales. La mayoría, si no todos, los operones de una célula bacteriana pertenecen al nivel más alto organizaciones reguladoras que nosotros llamaremos las REDES REGULADOR:
"El operon es un sello de la célula del procarioto. Este modo de organización - conque pueden unirse los genes de una senda entera como una sola unidad de las transcripciones - logra una solución sencilla al problema de genes del coregulato de función relacionada. ¿Por qué va más allá? ¿Hay una necesidad por la célula bacteriana, mover más allá de esta estrategia exitosa obvia?
"Hay dos respuestas por lo menos a estas preguntas. El primero es, que algunos procesos bacterianos involucran demasiados genes, ser acomodado en un solos operon laborables. Esta situación se ilustra por la maquinaria de traducción. Allí un grupo de por lo menos 150 productos del gen (el ribosoma ARN, las proteínas del ribosoma, la iniciación, el alargamiento, y la terminación factoriza, amino-acilo-tARN los sintetase, y tARNs) está directamente envuelto haciendo la proteína de los aminoácidos.
"Ellos son tan numerosos, que para coordinar su síntesis uniéndose sus genes en un solo operon, sería torpe, sino imposible. Todavía, la regulación coordinada de estos genes es importante a la eficacia global de crecimiento bacteriano. ... Esta coordinación es cumplida, aunque estos genes son organizados en muchas docenas de operons no-unión.
"La segunda respuesta es, que algunos procesos bacterianos involucran varios genes que deben ser independientemente ambos regulados y sujeto a un atropellar, coordinando el mando. Esta situación se ilustra el más claramente por el conjunto de genes, mientras poniendo en código enzimas catabólicas que están envuelto en la utilización de azúcares, aminoácidos, y otros compuestos para el carbono y energía. Durante el crecimiento en un ambiente que contiene una mezcla de cosas así compone, la economía exigiría, que sólo un substrato del premio que es el mejor capaz satisfaga el carbono y la energía necesita de la célula, debe metabolizarse. (Para las bacterias entericas, la glucosa es semejante substrato; para el pseudomonades, el succinate es.) Deben reprimirse los operons que ponen en código las enzimas para el metabolismo de los substratos secundarios, redundantes. Cada operon deben, sin embargo, se induzca individualmente, cuando su substrato del cognado está presente en la ausencia del substrato del premio. Este llamadas de requisito dobles para un nivel de organización sobre el del operon." (1990:351, 352).
¿Cuántos sistemas del multigene están allí en una célula bacteriana? ¿Cómo ellos trabajan? ¿Y cómo complejos ellos están?
Prof. F. C. Neidhardt y co-obreros: "Se estima, que la célula bacteriana ha desenvuelto varios cientos sistemas del multigene. Su descubrimiento y el análisis tiene sólo empezado. Algunos de los ejemplos bueno-conocidos para la bacteria enterica Escherichia coli son:
· Redes que están envuelto en la contestación a la limitación de uno u otro nutriente - el carbono y fuentes de energía, el amoníaco, el fosfato inorgánico,
· Redes que están envuelto en las reacciones de la oxidación-reducción y transporte del electrón
· Redes que se preocupan por la contestación dañar por la oxidación, radiación, temperatura alta y baja, y extremes de presión osmótica
· Las redes misceláneas, alguno con subredes que provoca los cambios del comandante en la fisiología y morfología de la célula.
La célula bacteriana ha desenvuelto las maneras diversas, para tejer el operons individual en las redes coordinadas; los varios mecanismos están empezando a ser elucidados simplemente. En algunos casos, el legado de un regulador de la proteína allosterico se ha pedido prestado simplemente de la regulación del operon: un repressor de la proteína o el activador reconoce una sucesión particular que es común a las regiones controlando del operons del miembro. Este legado se usa en el SOS, daño de la oxidación, y los sistemas de transporte de electrón anaerobios en las bacterias entericas.
"En otros sistemas la red se define por un factor de la sigma alternativo que reprograma el polimerase de ARN para reconocer a los promotores de la operons del miembro. El susto de calor y sistemas del sporulación de varias especies bacterianas ilustran esta situación. Otras redes involucran una combinación de reguladores de la proteína y sigma factoriza, como encuentre en el sistema de utilización de nitrógeno de muchas bacterias. Una de las redes más penetrantes, el operons severo, es regulado por el tetrafosfato de guanosine de nucleótido (el ppGpp) de una manera que todavía será elucidada.
"Porque se reconocieron muchas redes regulador primero como las contestaciones celulares a los cambios medioambientales, la estructura formal de un sistema de la estímulo-contestación puede mantener una función de la organización útil la masa de información sobre cada red. Un sistema de la estímulo-contestación (Figura 1 en su libro) contiene una senda que empieza con un estímulo del ambiente que afecta algún blanco celular o sensor que genera un signo. Este sistema de la estímulo-contestación directamente - o indirectamente, cuando los pasos señalados a través de uno o más transductores - afecta la actividad o síntesis de un regulador que controla el rendimiento. Ésta normalmente es alguna contestación adaptable al cambio medioambiental. Este sistema incluye a menudo un mecanismo de mando de regeneración que permite un retorno a la condición del prestimulus o a una nueva consonante de equilibrio con un ambiente cambiado.
"En una red del gen bacteriana, un estímulo del ambiente (por ejemplo, temperatura cambiada, estado nutriente, o toxicidad) se pintaría como afectar algún sensor que generaría un signo para afectar la actividad o síntesis de (normalmente) un regulador de la proteína de los genes del miembro. Los productos de los genes del miembro realizarían alguna función(s celular), para promover el crecimiento o supervivencia de la célula. Los modulador-demoduladores de alguna clase permitirían un retorno al estado del prestimulus de expresión del operons del miembro o establecerían un nuevo nivel de su expresión.
"Está claro... tantos del operons de E. coli ya se asignan los a uno o más redes. Con la ayuda del armazón, con tal de que en Figura 1, nosotros demostraremos unos de estas redes a fondo, para ilustrar principios de fisiología del multigene y la biología molecular. Las primeras dos redes, ser discutido, han sido en parte escogidas, debido a su importancia cuantitativa en la fisiología celular. Juntos ellos controlan directamente o indirectamente probablemente tres-cuarto de la capacidad proteína-sintetizando de la célula bacteriana." Neidhardt, F. C. (1990:352-357).
La represión del catabolito de alimentar los genes: ¿Lo que hace esa media? ¿Y cómo hace ese trabajo?
Prof. F. C. Neidhardt y co-obreros: "La Mayoría de las especies de destacamentos de la bacteria que un repertorio extenso de genes que ponen en código las enzimas catabolicas dirigió contra las docenas de compuestos diferentes de valor nutritivo diverso. El crecimiento en una sola fuente de carbono y energía - un substrato - requiere un nivel celular relativamente alto de las enzimas que metabolizan el substrato y alimentaban los productos catabólicos en las sendas alimentando centrales. Porque las sendas todas metabólicas en el flujo celular del metabolitos que se ha producido por estas enzimas catabólicas.
"Las bacterias aparecen darse cuenta de la adecuación de cada senda catabólica en una circunstancia dada y regular la expresión de flujo de gen de acuerdo con. Nosotros ya hemos notado, que el programa por decidir, qué enzimas para hacer y en qué cantidades, debe sofisticarse más que uno, qué simplemente requiere la célula, inducir las enzimas catabólicas para substratos que se ha dado cuenta de en el ambiente. De hecho, a pesar de un medio-siglo de esfuerzo intensivo, la sofisticación de este programa celular ha excedido la habilidad de fisiólogos, entender sus complejidades.
"Las bacterias entericas emplean cuatro procesos diferentes, asegurar el estado de prioridad de glucosa como un carbono y fuente de energía, por lo menos:
· La síntesis constitutiva: Las enzimas de catabolismo de glucosa, están en la medida grande una parte del EMP la senda alimentando central. Ellos se producen a los niveles altos, no importa qué substratos están presentes en el ambiente.
· La exclusión del inducido: la glucosa previene la entrada de ciertos otros substratos induciendo por la inactivación de su permeases.
· La represión transeúnte: la Suma de glucosa produce a un transeúnte, inhibición severa de síntesis de inducible las enzimas catabólicas para cerca la mitad de una generación a pesar de la presencia de su inducido.
· La represión catabólica: el Metabolismo de glucosa produce una inhibición continuada, pero menos severa de síntesis de la enzima inducido, incluso en la presencia de los inducidos apropiados.
"Se entienden Non de estos procesos totalmente. Y la represión del catabólito es particularmente compleja. Incluye un elemento, sin embargo sobre que mucho ha sido sabio: la CAMP CAP NETWORK. En E. coli el operon del lac es un miembro de esta red. El número total de operons del miembro no es conocido; pero pueden ser varios docena y puede incluir aquéllos, qué se preocupa por el metabolismo de galactose, arabinose, maltose, triptofan, D-serine, e histidine. Los operons del miembro poseen una sucesión del nucleótido característica a que la CAP de la proteína regulador (la proteína de activador de gen catabólica, producto del gen del crp) los lazos." Neidhardt, F. C. et al. (1990:357, 358).
¿El transducción señalado en los sistemas del multigene, lo que hace esa media? ¿Cómo eso trabaja? ¿Y cómo complejo está?
Prof. F. C. Neidhardt y co-obreros: "La red del arc con las otras tres redes, sólo describió, comparta un tema común: información que procesa por la interacción de la proteína-proteína. Permítanos considerar este tema en la profundidad mayor.
"El proceso de información fisiológica no es un nuevo tema para nosotros. Nosotros encontramos algunos aspectos de información procesar, cuando nosotros consideramos los cambios del conformacional en cierto (allosterico) proteínas que se provocan por el complejo reversible de moléculas pequeñas específicas. Las moléculas pequeñas - qué es productos del extremo de sendas del biosintetico, induciro de sendas catabólicas, metabolitos del precursor, o nucleótidos - llevó la información (sobre su concentración celular) a la proteína allosterica. Este proceso consideró para la mayoría del mando en la actividad de la enzima en las sendas del biosintetico y alimentando las reacciones. Y también para el dispositivo estratégico central de fisiología del operon - la activación y represión de transcripción por proteínas que en una estructura pueden ligar ADN y en otro, no pueda." (1990:375-377).
¿Cómo la célula bacteriana regula sus redes del multigene? ¿Por qué estas interacciones son importantes?
Prof. F. C. Neidhardt y co-obreros: "Las interacciones de la Proteína-proteína son importantes regulando las redes, describió anteriormente. Pero ellos no son los solos medios de regular las redes del multigene. De hecho, los medios moleculares por que los grupos de operons diferente son coordinados en su expressio, son bastante diversos.
"Nosotros consideramos que tres redes que se parecen entre sí en un aspecto funcional para ilustran esta diversidad. Cada uno se induce a la expresión de alto nivel como el resultado de un solo, bien-definido cambio del parámetro medioambiental. Pero el mecanismo de la inducción es muy diferente en cada caso.
"Un cambio a la temperatura alta induce un juego de cerca 20 genes en la mayoría de las células bacterianas. Esta inducción constituye la contestación del calor-susto. Es a una magnitud increíble conservada del arquebacteria y eubacteria a través del más bajo eucariotes a las plantas más altas y animales. Virtualmente cada célula microbiana y cada planta y la célula animal responden a un cambio-despierto en la temperatura. A un rango que simplemente es anterior que es normal para el organismo dado induciendo la síntesis de sus proteínas del calor-susto.
"Otra red - con cerca el mismo número de genes - es el sistema de SOS. Este grupo de genes se induce a la expresión de alto nivel, cuando el daño de experiencias celular a es ADN, como por la irradiación de UV. Los productos de la proteína de estos genes están envueltos en los varios procesos, ese daño de la reparación a ADN, y ese impide a las células dividir, hasta que él tal daño se repare. Esta red es inducido por una hendidura proteolitica del repressor del operons del miembro.
"El repressor es el producto del gen del lexA; en el daño al ADN de una célula. La proteína de RecA se activa, para facilitar la hendidura proteolitica de LexA. Induce los genes LexA-controlados que constituyen el sistema de SOS por eso.
"Todavía otro modo se ilustra por la red del oxyR. Este grupo de una docena o para que los genes sean inducidos por el oxidative dañe por que se trae adelante por ejemplo, peróxido de hidrógeno. Los genes del miembro ponen en código las enzimas proteccionista, como el catalase-hydroperoxidase yo y alkylhidroxiperoxidase. Esta red es inducido por la activación del regulador positivo OxyR, el producto del gen del oxyR.
"Estos ejemplos sirven, para ilustrar el hecho que la regulación de redes del gen es como diverso, como es la regulación de operons individual." Neidhardt, F. C. (1990:380, 381).
¿Por qué hace las proteínas de la hechura celulares vivientes? ¿Y por qué sabe, cómo hacerlos? ¿Y por qué un cierto juego de ácidos nucleicos traduce el texto en las proteínas? ¿En otros términos: Por qué hace una cierta señal (o carta) del ADN codifique sepa, lo que significa? ¿Y por qué significa esto?
Bruce Alberts y co-obreros admiten en su libro de texto la Molecular Biology of the Cell (Biología Molecular de la Célula) (1989:219), aunque ellos creen en la evolución: "Los procesos moleculares en que la síntesis de la proteína es basada, parezcan inexplicablemente complejos. Aunque nosotros podemos describir muchos de ellos, ellos no tienen el sentido conceptual de la manera que la trascripción de ADN, reparación de ADN, y repetición de ADN hacen. Cuando nosotros hemos visto, la síntesis de la proteína en los centros de los organismos actuales en una máquina del ribonucleoproteína muy grande, el ribosoma. Consiste en proteínas que se colocan alrededor de un centro de moléculas del rARN.
"¿Por qué las moléculas del rARN deben existir en absoluto? ¿Y cómo ellos vinieron a tocar semejante parte dominante en la estructura y función del ribosoma? La respuesta nos ayudaría, entender la síntesis de la proteína, indudablemente. ... La síntesis de la proteína también confía pesadamente en un número grande de proteínas diferentes que se ligan al rARNs en un ribosoma. La complejidad de un proceso, con los tantos componentes entrelazados diferentes, ha hecho a los muchos biólogos desesperar. Porque ellos no creen, que ellos alguna vez podrán entender la senda por que la síntesis de la proteína evolucionó."
Nosotros todavía podríamos recordar, lo que Profesor Bruce Alberts y sus co-obreros exigieron en su libro de texto The Cell (La Célula) (1989:41): "No hay ningún cuarto en la vista contemporánea de vida... para nada más fuera de las leyes de química y físicas."
¿Es ese lógico, es ese razonable, es ese científico? Nada. La declaración: "No hay ningún cuarto en la vista contemporánea de vida para algo fuera de las leyes de química y físicas." No es lógico, no es razonable, no es científico. La razón: UNA ley siempre viene de un dar de ley. Las leyes de química y física contienen información científica más alta y matemática. Los científicos humanos abajo aquí en la tierra han entendido ahora hasta sólo unos de ellos. Algunas de estas leyes de química y físicas ellos han podido sólo entender un poco el pedazo. La información y matemática existen independientemente de humanidad. El científico humano sólo ha encontrado y los ha entendido un poco. La información y matemática son algo el no-material, espiritual. Ellos tienen su fuente en un espiritual, el mundo del no-material, en el verdadero Dios de la Biblia cuyo nombre es Jehová.
Ya en el nivel del código nosotros podemos saber, que se ha pensado fuera y se ha hecho por una persona inteligente. Un código (o alfabeto) es algo espiritual, el no-material. Una persona inteligente debe pensarlo primero fuera. Él debe seleccionar ciertas señales primero (las letras, etc.) y decide, lo que ellos querrán decir. Con estas señales, él puede escribir las palabras, frases, capítulos, y el libro entero entonces.
· Los 20 aminoácidos son las 20 letras del código del aminoácido. Con estas 20 letras (y la gramática proporcionó con él), las hechuras celulares vivientes (escribe) sus proteínas.
· Los 4 ácidos nucleicos son las 4 letras del código ácido nucleico. Con estas 4 letras (y la gramática proporcionó con él), las hechuras celulares vivientes (escribe) su código genético, su cadena de ADN.
La primera célula del arquea en la tierra era así como complejo como aquellos, qué está viviendo ahora. No tiene "descubrió" o "inventó" algo. Incluso el más "sencillo" el organismo del una-célula es más complejo, que algo, qué hombre tiene puede hacer cultive ahora. El excelente-computadora más rápido de hoy es simplemente el juguete de un niño primitivo, comparó a la célula más sencilla que está viviendo hoy y qué se ha mantenido en esta tierra, hace unos 3.8-4.0 mil millones años, al principio.
La hipótesis de evolución, o neo-Darwinismo, como ahora normalmente enseñó a lo largo del mundo, no tiene nada que hacer en absoluto con la ciencia seria. Es una religión falsa. Tiene sus raíces en las enseñanzas de Babilonia antigua (Sumer-Accad) y Egipto. Sus sacerdotes paganos ya enseñaron hace unos 3.500 a 4000 años, esa vida en la tierra ha evolucionado solo del agua y barro del Éufrates y Tigres y del Nilo. La hipótesis moderna de evolución o neo-Darwinismo es un fraude grande. Esconde su verdadero significado detrás del vestido blanco de ciencia moderna.