Capítulo 6: La Pared Celular
Las bacterias Gram-positivas y Gram-negativas. Su membrana y muro. Cómo ellos hacen sus paredes. Cómo ellos difieren. ¿Qué fue necesitado, pensarlos fuera y hacerlos? ¿Su pared celular demuestra, que ellos todos han venido de un antepasado común?
Prof. Frédéric C. Neidhardt y co-obreros escriben en su libro de texto Physiology of the Bacterial Cell (Fisiología del de la Célula Bacteriana) (1990:25-27) sobre la pared de las bacterias: "Las Bacterias pueden ser divididas en dos clases anchas - Gram-positivo y Gram-negativo. Las bacterias Gram-positivas y Gram-negativas difieren en varios aspectos importantes, además de la estructura del sobre. La distinción Gram-positivo-Gram-negativa es una profunda. No sólo refleja una diferencia fundamental en la estructura de la pared, pero otras diferencias bioquímicas, fisiológicas, y genéticas importantes también. La mayoría, pero no todos, las bacterias, son miembros de estos dos grupos. Esos ese otoño fuera de estos grupos es de dos clases. Un grupo es estrechamente similar a otras bacterias, pero completamente falta las paredes - el Mollicutes. El otro grupo, las Arquebacterias, produce paredes que no contienen un mureino típico y en muchos otros respetos difiera de la célula del procariota típica. (1990:25-27).
La Pared celular. ¿Qué científicos han averiguado ahora sobre la pared celular? ¿Cómo complejo está? ¿Qué debe uno que todos saben en el orden entender y hacerlo?
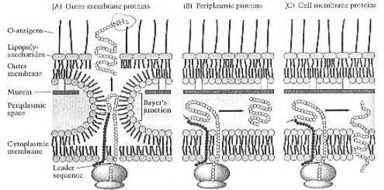
Prof. F. C. Neidhardt y co-obreros: "Como las células van, las bacterias son particularmente duras; es decir, ellos son difíciles romper por los medios mecánicos. Y ellos retienen su forma incluso bajo las condiciones ásperas. Mucha de la dureza de bacterias se relaciona a su pared celular que proporciona el apoyo mecánico rígido e impide a la presión estallar las células (él lisies osmótico). Además, la pared celular representa a un químico y la defensa física contra químicos nocivos que pueden dañar la membrana celular. Las bacterias Gram-positivas y Gram-negativas difieren considerable en la estructura de sus paredes celulares, y en el papel de esta estructura, cubriendo con los cambios medioambientales. Cualquier porción de la pared de miradas de las células Gram-positivas como una manta espesa, el de la mayoría de las células Gram-negativas como una hoja débil. Incluso en la realidad, la pared Gram-negativa delgada tiene la fuerza tensor considerable." (1990:33).
La Pared Gram-positiva. Prof. F. C. Neidhardt y co-obreros: "La pared celular Gram-positiva consiste en una chaqueta del multimolecular espesa de muerein (un tipo de peptidoglican) con las cantidades menores de otros polímeros, notablemente los ácidos del teichoicos; esparció. El tejido polimero consiste en muchas capas que se envuelven alrededor de la longitud y anchura de la célula. Por eso ellos forman una bolsa que determina el tamaño y forma del organismo. El mureino puede aislarse intacto como una estructura, llamó el sacculus del mureino. La forma de la bacteria depende de la forma del sacculus. La mayoría de la mirada de la bacteria como las varas (las bacili), o hélices (las spirochetes); unos mirada como los husos, estrella de mar, o el polihedra chato; todavía otros asumen las formas más complejas, diferenciadas. ... La red del mureino rígido permite bacterias, sobrevivir en medios de comunicación que normalmente son los hipotonico. Es decir, ellos tienen un más bajo presión osmótica, que el de su citoplasma. En la ausencia de un rígido para empujar contra, la membrana estallaría corsé-como la estructura, y el lise de las células (= fluya separadamente)." (1990:33-36).
La Pared Celular Gram-negativa y la Membrana exterior. Prof. F. C. Neidhardt y co-obreros: "Las bacterias Gram-negativas han desenvuelto una solución sumamente diferente al problema de proteger la membrana citoplasmica. Su capa del mureino está muy más delgada, que el de bacterias Gram-positivas, y ellos hacen una estructura completamente diferente - una membrana exterior que se construye a fuera de una capa del mureino delgada. La membrana exterior es químicamente distinta de las membranas biológicas usuales. Es capaz, resistirse los químicos perjudiciales. Es una estructura del bicapa, y su hoja impresa interna se parece en la composición que de la membrana citoplasmica.
"Su hoja impresa exterior, por otro lado, tiene un único elector en el lugar de fosfolipid. Este componente es el lipopolisacarido bacteriano, o LPS. Ésta es una molécula compleja que no se encuentra en otra parte en la naturaleza. Como resultadas, las hojas impresas de esta membrana son sumamente asimétricas. Y las propiedades de este bicapa raro difieren considerablemente de aquellos de una membrana biológica regular. La habilidad, excluir los compuestos hidrófobos, es rara entre las membranas biológicas.
"La exclusión de compuestos hidrófobos en las bacterias Gram-negativas, como en las bacterias Gram-positivas, es cumplida rodeando las células con los polisacáridos hidrófobos. Pero éstos difieren en la estructura y organización en los dos grupos. La membrana exterior, sin embargo, los regalos un callejón sin salida claro: puede esperarse que excluya los compuestos hidrófilos también debido a su naturaleza del lípido. En ese caso, ningún compuesto, hidrófobo o hidrófilo, podría cruzar la membrana exterior.
"Mientras resolviendo el problema de protección de la membrana citoplasmica, las bacterias Gram-negativas aparecen haber creado un nuevo. ¿Cómo estos organismos transportan sus nutrientes? ¿Ellos copian en la membrana exterior los dispositivos de transporte activos de la membrana citoplasmica? Esta estrategia no sólo sería una inversión malgastadora, pero también podría hacer la membrana exterior así como sensible a los desafíos medioambientales, cuando la membrana citoplasmica es. Una vez más, las bacterias han encontrado una solución interesante: La membrana exterior tiene los cauces especiales, ese permiso la difusión pasiva de compuestos hidrófilos como los azúcares, aminoácidos, y ciertos iones. Estos cauces consisten en moléculas de la proteína, los porinos inclinadamente llamados. Los cauces del porin son estrechos, sólo el tamaño correcto, para permitir la entrada de compuestos más pequeño que 600-700 dalton (Figura 9 en su libro).
"Los cauces son tan pequeños, que los compuestos hidrófobos entrarían en el contacto con el polar 'la pared' del cauce y por eso se excluya. A alguna magnitud, E. coli pueden seleccionar los porinos con los tamaños del cauce diferentes, en la contestación a las propiedades osmóticas del medio.
"Ciertos compuestos hidrófilos que las necesidades celulares para sobrevivir, son más grandes, que la exclusión de porin. Estas moléculas grandes incluyen la vitamina B12, azúcares que son más grandes que el trisaccarides y el celates férrico. Cosas así componen la cruz la membrana exterior por los mecanismos de la infiltración separados, específicos. Ellos utilizan proteínas que se han diseñado sobre todo al translocate cada uno de estos compuestos. Así, la membrana exterior permite el pasaje de compuestos hidrófilos pequeños, excluye compuestos hidrófobos, grande o pequeño, y permite la entrada de algunas moléculas hidrófilas más grandes por los mecanismos sobre todo diseñados." (1990:37-40).
"La membrana exterior no es una estructura completamente separada; se conecta a la capa del mureino y la membrana citoplasma. La conexión con la capa del mureino se media por dos tipos de interacciones, el más importante es un lipoproteína de la membrana exterior. Esta proteína está presente en unas 700,000 copias por célula que le hace la proteína más abundante (numéricamente, no por el peso) en E coli.
"Sobre un tercio de las moléculas del lipoproteína covalente sé unidos juntos al mureino y sostenimiento de ayuda las dos estructuras están. El otro tipo de interacción es el firme (pero probablemente no el covalente) la asociación de algunos de los porinos de la membrana exteriores con el mureino. Juntos, los lipoproteínas y porinos proporcionan en cada célula encima de 400,000 puntal-como avisas entre la membrana exterior y la pared celular." (1990:41).
¿Qué substratos específicos son la bacteria E. coli capaz para subir? ¿Cómo los sube, de qué maneras diferentes?
Prof. F. C. Neidhardt y co-obreros: "De los varios mecanismos de transporte, casi todos (incluyendo difusión facilitada, transporte activo susto-sensible, y el transporte activo secundario) participe en la captación de uno u otro azúcar o alcohol de azúcar por E. coli (Figura 4 en su libro). Interesantemente, la captación de un azúcar particular no sigue un modelo: por ejemplo se toma la lactosa en E. coli por el transporte del protón, pero en Staphylococcus aureus por unas PTS.
"Los aminoácidos son transportados por los sistemas de transporte activos susto-sensibles o secundarios. En E. coli, 14 sistemas de transporte diferentes que traen los aminoácidos en la célula. De un examen de los sistemas de transporte de aminoácido de bacterias, varios modelos surgen. Alguno del transporte de los sistemas un grupo de aminoácidos con la estructura similar. A menudo éstos tienen los subsistemas para único de los aminoácidos. Esta redundancia clara sirve un propósito. Uno de los sistemas tiene la afinidad alta (la afinidad 10 veces constantes o más mayor) y el flujo alto. Cada uno tienen las ventajas obvias en particular los ambientes. Además de los sistemas de transporte de aminoácido, E. coli tiene una variedad de sistemas de transporte por los cofactores, varios iones, y los intermedios metabólicos." (1990:182, 183).
Cómo una Gramo-negativo-bacteria hace sus propias proteínas del sobre. De Neidhardt et al. de Fig. 8 del (1990:114). La célula minúscula ha alcanzado aquí, qué hombre todavía no puede hacer al principio de este siglo XXI: montaje automático, producción automática. El, que ha pensado hacia fuera y ha hecho este unicélule organismo, debe saber tan mucho más sobre ciencia y tecnología, que cualquier científico ahora que vive en esta tierra: el creador, que nombre es Jehová.
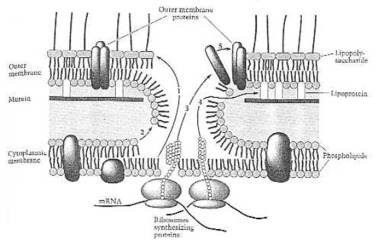
Reacción de la asamblea de la membrana externa. (1) Lipopolysaccharide; (2) phospholipids; (3) proteínas externas de la membrana; (4) lipoproteína; (5) uno mismo-montaje de la membrana externa. De Neidhardt et de Fig. 14 del al. (1990:121).
¿Cómo hace E. coli superado él extreme de presión osmótica medioambiental? ¿Y cómo hace sus porins, los poros en su membrana exterior?
Prof. F. C. Neidhardt y co-obreros: "Una variable medioambiental importante de que las bacterias deben tratarse, es el osmolaritía de su ambiente. La célula bacteriana utiliza varias estrategias moleculares, neutralizar los efectos de extremes deletéreo de presión osmótica medioambiental.
"La membrana exterior se cruza por cauces que están rayado por las proteínas los porinos llamados. En E. coli hay dos porinos mayores - OmpF y OmpC. Ellos cuidan ser hechos como las alternativas a nosotros. Los poros hechos por OmpC son ligeramente más pequeños y son preferencialmente hecho a la presión osmótica más alta y la temperatura más alta. Este modelo significa, ese OmpC es la forma predominante durante el crecimiento de bacterias entericas en el tracto intestinal. Tiene el sentido fisiológico bueno para las bacterias entericas tener los poros más pequeños, al residir en el tracto intestinal y el más grande fuera. Los poros más pequeños excluyen muchas de las moléculas tóxicas encontrados en el tracto intestinal. Los poros más grandes permiten los substratos de que diluye solución, entrar en el poro celular prontamente, porque los solutes deben difundir a través de los poros de la membrana exteriores. Y la proporción de difusión es proporcional al área cruz-particular del poro, así como la diferencia de la concentración por la membrana.
"Los genes que ponen en código los porinos mayores así como ciertos otros genes que afectan las propiedades de la membrana exteriores son controlados por dos proteínas - los productos del envZ unido (para las proteínas del sobre) y ompR (para la regulación de proteína de membrana exterior) los genes. La proteína de EnvZ es una proteína de la membrana citoplasmica que se piensa para funcionar como un osmo-sensor. Tiene la similitud estructural fuerte a cierto kinases de la proteína; sobre la base de a esta similitud y otra evidencia, y del fenotipo de cosa sin sentido y otras mutaciones en el envZ, investigadores creen, ese EnvZ actúa recíprocamente con la segunda proteína, OmpR. Afecta su actividad, por el fosforilato él.
"La proteína OmpR liga el specifical al río arriba de ADN de promotores que controla; el sitio obligatorio está alrededor de -50 en el promotor del ompF, y -90 en el promotor del ompC. Un modelo actual, para cómo la expresión alternativa de ompF es cumplida, es como sigue (Figura 7 en su libro): la presión osmótica alta lleva al fosforilación de EnvZ. Y esto activó la proteína a su vez el fosforilates OmpR. Fosfo-OmpR (OmpR-P) reprime trascripción de ompF, ligando a un sitio entre -40 y -60 en la región de promotor de ompF; activa transcripción del ompC, ligando a los sitios de bajo-afinidad entre -75 y -105 en la región de promotor de ompC. Cuando la presión osmótica es baja, el nivel de OmpR-P es bajo. Y por consiguiente es incapaz, para activar el ompC; pero todavía puede activar el ompF, ligando a los sitios de alto-afinidad en la -60 a -100 región del promotor del ompF.
"Los efectos de OmpR-P, como un stimulator y como el inhibidor de trascripción de estos dos genes, se complementa por otro mecanismo. El micF del gen se transcribe en la dirección opuesta del ompC de un promotor, cerca del de ompC; el micF pone en código un 174-base ARN que es complementario a una región cerca de la salida del mensaje del ompF. La trascripción del micF se ha mostrado, para bloquear expresión de ompC. Porque la regulación de expresión del micF es similar a eso de ompC, las condiciones que inducen el ompC inducirán el micF simultáneamente, y por eso las ayudas reprimen el ompF (Figura 7 en su libro).
"Hay una red de genes además de aquellos para los dos porinos que se regulan por el par del envZ/OmpR de proteínas entrelazadas; algunos de estos genes son inducidos, otros reprimieron. El sentido fisiológico global de la opción de genes que son incluido en este sistema todavía es huidizo; la importancia a nosotros de la historia de EnV/OmpR es, que ha proporcionado un modelo importante de interacción de la proteína-proteína en la estímulo-contestación que conecta una red de computadoras, mientras incluyendo muchos para que la función fisiológica sea de cristal claro." Neidhardt, R. C. et al. (1990:272, 373).
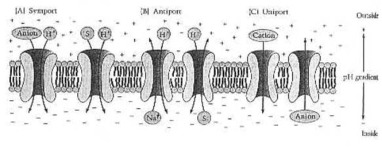
Sistemas secundarios del transporte activo. (a) Reacciones de Symport. El gradiente del pH creado por las impulsiones del transporte activas primarias (a la izquierda) un symport electroneutro de un anión con un protón, y (a la derecha) un symport electrogénico de un solute uncharged (s) junto con un protón. (b) Reacción de Antiport. El gradiente del pH conduce (a la izquierda) el intercambio electroneutro de un catión para un protón y (a la derecha) el intercambio electrónico de un solute encarga (s) para un protón. (c) Reacciones de Uniport. El gradiente del pH conduce un catión en la llamada (izquierda) de un anión hacia fuera (derecho). De Neidhardt et al. de Fig. 1 (1990:178).
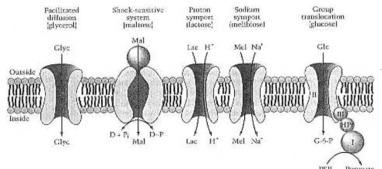
Mecanismos portador-mediados para el entrada (uptake) de azúcares y de alcoholes de azúcar por E. coly. El glicerol (Glyc) entra por la difusión facilitada. La maltosa (Mal) entra por un dependiente dar una sacudida eléctrica-sensible del sistema en una proteína obligatoria (esfera), dos proteínas del transmembran (óvalos), y una proteína de la energía-transaducho (flecha curvada, doble-dirigida) que hidrolice a donante de gran energía del fosfato (D~P). La lactosa (Lc) entra por el symport y el melibiose (mel) de la proteína por un symport de Na+ del transporte activo secundario. La glucosa (glc) entra en por un PTS con la mediación de enzimas I (i), HPr, la enzima II (ii), y la enzima III (iii); el producto intracelular del proceso es la glucosa 6-phosphate (G-6-p). De Neidhardt et del al. (1990:182) Fig.4. - si usted, lector querido, realmente no ha entendido esto, no preocuparse. Acabo de desear demostrarle, usted complicó esto es, y cuánto el, que ha pensado el suyo hacia fuera, tiene que saber sobre ciencia natural, alrededor, la física y la bioquímica.
...
Primero la Membrana, cómo se levantado
¿Cómo la primera membrana ha entrado en ser? ¿Podría tener el airsen solo en la sopa química primordial?
Prof. Siegried Scherer: "Fosfolipides son un principal-componente de los bio-membrana. Ellos son hechos de la glicerina, ácido fosfórico y compuestos de alifato de largo-cadena. Ellos se unen junto con la glicerina como éteres o esters. Estas moléculas, como todas las otras substancias que están activo en la superficie (el tenside) tenga un hidrófobo (= agua-evitando), y un hidrófilo (= agua-amoroso), componente. Las moléculas con estas estructura-características son capaces, venir juntos espontáneamente. Por ejemplo, como las doble-capas, micélula, y vesículas). Cómo las tales substancias pudieran levantarse bajo las condiciones prebioticas, no es conocido.
"Las membranas biológicamente activas, cuando nosotros los conocemos de las células vivientes, siempre origine de ya membranas existentes. Ellos no se sintetizan del novo (encima de de nuevo, nuevamente). El bio-membrana no sólo debe demarcar la célula contra sus ambientes. Pero también debe, entre otras cosas, material de transporte a través de su membrana. La construcción compleja de espejos de los biomembrana las tareas diversas de las membranas celulares. Por ejemplo, mecanismo de reconocimiento, célula-célula-comunicación)." (1998:146).
Hans Kleinig está un profesor para la célula-biología en la Universidad de Friburgo. Uwe Maier está un profesor para la célula-biología en la Philipps-Uuniversidad Marburg, Alemania. Ellos escriben en su libro de texto del Zell Biologie (Célula-biología) (1999:73) sobre la biosíntesis de membranas: "Los Biomembrana no pueden levantarse del novo en el citoplasma de una célula. Los membrana-componentes, normalmente se sintetizan proteínas, como los lípidos, en las membranas ellos: en el protocites: en la plasma-membrana..."
Las membranas biológicamente activas, cuando nosotros los conocemos de las células vivientes, siempre origine de ya membranas existentes. Ellos no se sintetizan nuevamente. Cómo las tales substancias pudieran levantarse bajo las condiciones prebióticas, no es conocido
Nosotros hemos mirado ahora brevemente células Gram-positivas y las células Gram-negativas. La célula Gram-positiva tiene una pared. La célula Gram-negativa tiene una membrana. Las células Gram-positivas, células Gram-negativas, y arquebacterias difieren mucho de nosotros. Ellos no se relacionan en absoluto a nosotros. Ellos son así como diferente de nosotros, como ellos son de las plantas y animales. Y uno no puede rastrearlos atrás a un antepasado común mítico. Cerca del principio de la historia de tierra, había células Gram-positivas y Gram-negativas ya hace unos 3.5 a 3.8 mil millones años, y arquebacterias, sólo gusta hoy.
Muchas del genera y especies, identificados a Warrawoona, el NO Australia, todavía está viviendo hoy. Éstos que los organismos del una-célula eran así como complejo, como esos viviente hoy. Ellos no han evolucionado en absoluto en las formas más altas de vida. Ellos son fábricas químicas totalmente automáticas, con un diámetro de un fragmento diminuto de un milímetro. ¿Cómo haga las fábricas químicas, las casas, con sus paredes y tejados, se levantan hoy? Una casa (o la fábrica química) se levanta al principio en la mente de un ser humano inteligente.
· Primero, él tiene una necesidad. Él podría decirle: "Yo quiero a él haga caluroso y seca, dónde los osos y lobos no pueden venir y pueden matarme."
· Entonces él tiene un deseo: "Yo quiero tener una casa (o una tienda, o una cueva)."
· Entonces él hace un plan: "Mi casa será fulano de tal grande. Tendrá fulano de tal muchos cuartos, puertas, y ventanas. Y yo lo haré con los ladrillos (o piedras, o leños)."
· Entonces él usa su plan de la construcción (el cianotipo), energía, y construyendo los materiales, y hace la casa.
Los primeros tres pasos que llevan a la casa son completamente espirituales, el no-material. Un ser humano inteligente, especializado como un constructor de la casa, los ha pensado primero fuera. Incluso los ladrillos en la casa (su forma y función), un ser humano inteligente ha pensado primero fuera. Los átomos que constituyen los ladrillos (las piedras, y leños), no sepa nada sobre una casa.
Materialistas han exigido: Todo viene de la materia. Pero una casa no crece a fuera de las piedras o leños de la taiga Siberiano, como un árbol hace. ¿Por qué no? Porque el material del edificio de la casa no sabe nada sobre la forma y función de una casa. Todos que vienen primero de un ser humano inteligente que ha pensado sobre esto.
Los átomos de la pared (o membrana) de la bacteria están a su nivel orgánico más bajo. Ellos son sólo un medios a un extremo, como los ladrillos en la casa. Una bacteria es mucho más complicada, que cualquier casa que el hombre ha construido alguna vez. El hombre puede construir una casa (y una fábrica química). Pero él no es capaz, hacer la célula viviente, una fábrica del químico automatizada llena. Es demasiado complicado. Es más complicado, que algo, qué hombre ha hecho en la vida ahora hasta. Las primeras células vivientes en la tierra, hacen unos 3.5-4 mil millones años, eran así como complicado, como esos viviente hoy.
La célula contiene la habilidad bioquímica alta. Contiene información y matemática. Pero la propia célula no ha descubierto o los ha inventado. La información y matemática son algo el no-material, espiritual. Ellos tienen su fuente en un espiritual, el mundo del no-material. De, también la bacteria se debe de haber pensado primero fuera y se debe de haber hecho por alguien, quién sabe mucho más sobre la ciencia, que cualquier ser humano, manteniéndose en esta tierra ahora: el Creador, el Dios de la Biblia cuyo nombre es Jehová.